Pamoka yra skirtas visų profilių mokymo, visų formų teorinio pasirengimo užsiėmimams, įskaitoms ir egzaminams krypties „Biologija“ studentams. Vadovas apima pagrindinius struktūrinės biochemijos skyrius: pagrindinių biologinių makromolekulių klasių struktūrą, fizikines-chemines savybes ir funkcijas. Daug dėmesio skiriama daugeliui taikomųjų biochemijos aspektų.
Nukleotidai ir nukleorūgštys
Nukleotidų ir azoto bazių sandara
Nukleotidai dalyvauja daugelyje biocheminių procesų, taip pat yra nukleorūgščių monomerai. Nukleino rūgštys užtikrina visus genetinius procesus. Kiekvienas nukleotidas sudarytas iš trijų tipų cheminių molekulių:
azoto bazė;
Monosacharidas;
1-3 fosforo rūgšties likučiai.
Skirtingai nuo monosacharidų, nukleotidai kaip monomerai yra sudėtingos molekulės, susidedančios iš struktūrų, priklausančių skirtingoms klasėms. cheminių medžiagų, todėl būtina atskirai atsižvelgti į šių komponentų savybes ir struktūrą.
Azoto bazės
Azoto bazės yra heterocikliniai junginiai. Be anglies atomų, heterociklo sudėtis apima azoto atomus. Visos azoto bazės, esančios nukleotiduose, priklauso dviem azoto bazių klasėms: purinui ir pirimidinui. Purino bazės yra purino dariniai – heterociklas, susidedantis iš dviejų ciklų, vienas penkių narių, antrasis šešių narių, numeracija tokia, kaip parodyta paveikslėlyje. Pirimidino bazės yra pirimidino dariniai ir susideda iš vieno šešių narių žiedo, numeracija taip pat parodyta paveikslėlyje (31 pav.). Pagrindinės pirimidino bazės tiek prokariotuose, tiek eukariotuose yra citozinas, timinas Ir uracilas. Dažniausiai pasitaikančios purino bazės yra adeninas Ir guaninas. Dar du – ksantinas Ir hipoksantinas– yra tarpiniai jų metabolizmo procesai. Žmonėms oksiduota purino bazė veikia kaip galutinis purino katabolizmo produktas. - šlapimo rūgštis. Be aukščiau paminėtų penkių pagrindinių bazių, žinomos ir rečiau atstovaujamos nedidelės bazės. Kai kurių jų yra tik bakterijų ir virusų nukleorūgštyse, tačiau daug jų yra ir pro- bei eukariotinėse DNR bei transportinėse ir ribosominėse RNR. Taigi tiek bakterijų DNR, tiek žmogaus DNR turi daug 5-metilcitozino; 5-hidroksimetilcitozinas rastas bakteriofaguose. Neįprastos bazės buvo aptiktos pasiuntinio RNR - N 6 -metiladenine, N 6 , N 6 -dimetiladenine ir N 7 -metilguanine. Bakterijos taip pat turi modifikuotą uracilą su (α-amino, α-karboksi)-propilo grupe, prijungta N3 padėtyje. Šių pakeistų purinų ir pirimidinų funkcijos nėra visiškai suprantamos, tačiau jie gali sudaryti nekanoninius ryšius tarp bazių (tai bus aptarta toliau), sudarydami antrines ir tretines nukleorūgščių struktūras.
31 pav. Azotinių bazių struktūra
Augalų ląstelėse rasta purino bazių su metilo pakaitalais serija. Daugelis jų yra farmakologiškai aktyvūs. Pavyzdžiui, kavos pupelės, kuriose yra kofeino (1,3,7-trimetilksantino), arbatos lapai, kurių sudėtyje yra teofilino (1,3-dimetilksantino), ir kakavos pupelės, kurių sudėtyje yra teobromino (3,7-dimetilksantinas).
purino ir pirimidino bazių izomerija ir fizikinės bei cheminės savybės
Azoto bazės molekulė sudaro kintamų viengubų ir dvigubų jungčių sistemą (konjuguotų dvigubų jungčių sistemą). Tokia organizacija sudaro standžią molekulę, be konformacinių perėjimų galimybės. Dėl to negalima kalbėti apie azotinių bazių konformacijos pasikeitimą.
Azoto bazių atveju buvo nustatytas tik vienas izomerijos tipas, keto-enolio perėjimas arba tautomerizmas.
Tautomerizmas
Dėl keto-enolio tautomerijos reiškinio nukleotidai gali egzistuoti laktimo arba laktamo pavidalu, o fiziologinėmis sąlygomis vyrauja guaninas ir timinas (32 pav.). Šios aplinkybės svarba paaiškės aptariant bazių poravimosi procesus.
32 pav. Nukleotidų tautomerija
Tirpumas
Esant neutraliam pH, guaninas yra mažiausiai tirpus. Kitas eilėje yra ksantinas. Uratų pavidalo šlapimo rūgštis santykinai tirpsta esant neutraliam pH, bet labai blogai tirpsta žemesnio pH skysčiuose, pavyzdžiui, šlapime. Guanino žmogaus šlapime paprastai nėra, o ksantinas ir šlapimo rūgštis yra įprasti jo komponentai. Paskutiniai du purinai dažnai randami šlapimo takų akmenyse.
šviesos sugertis
Dėl konjuguotų dvigubų jungčių sistemos visos azoto bazės sugeria ultravioletinėje spektro dalyje. Absorbcijos spektras – optinio tankio pasiskirstymo grafikas priklausomai nuo bangos ilgio. Kiekviena azotinė bazė turi savo sugerties spektrą, pagal jį galima atskirti įvairių azotinių bazių tirpalus ar junginius, kuriuose yra azotinė bazė (nukleotidai), tačiau visų sugerties maksimumas sutampa esant 260 nm bangos ilgiui. Tai leidžia lengvai ir greitai nustatyti tiek azoto bazių, tiek nukleotidų ir nukleorūgščių koncentraciją. Absorbcijos spektras taip pat priklauso nuo tirpalo pH (33 pav.).
33 pav. Įvairių azotinių bazių sugerties spektrai
Azotinių bazių funkcijos
Azoto bazių laisvoje būsenoje praktiškai nėra. Išimtis yra kai kurie alkaloidai ir šlapimo rūgštis.
Azoto bazės atlieka šias funkcijas:
Jie yra nukleotidų dalis;
Kai kurie alkaloidai yra azoto bazės, pavyzdžiui, kavoje esantis kofeinas arba arbatoje esantis teofelinas;
Tarpiniai azoto bazių ir nukleotidų apykaitos produktai;
Šlapimo rūgštis yra priežastis urolitiazė;
Iš kai kurių organizmų azotas išsiskiria šlapimo rūgšties pavidalu.
Nukleotidai ir nukleozidai
Nukleozidų molekulės sudaromos iš purino arba pirimidino bazės, prie kurios atitinkamai N 9 arba N 1 padėtyje (β jungtimi) yra prijungtas angliavandenis (dažniausiai D-ribozė arba 2-deoksiribozė). Taigi, adenino ribonukleozidas (adenozinas) susideda iš adenino ir D-ribozės, prijungtos N 9 padėtyje; guanozinas- iš guanino ir D-ribozės N 9 padėtyje; citidinas- iš citozino ir ribozės N 1 padėtyje; uridinas- iš uracilo ir ribozės N 1 padėtyje. Taigi purino nukleoziduose (nukleotiduose) azoto bazė ir cukrus yra sujungti 1-9 β glikozidine jungtimi, o pirimidinuose - 1-1 β glikozidine jungtimi.
2'-dezoksiribonukleozidų sudėtis apima purino arba pirimidino bazes ir 2'-dezoksiribozę, prijungtą prie tų pačių N 1 ir N 9 atomų. Ribozė arba 2′-dezoksiribozė prisijungia prie žiedinės bazės struktūros per santykinai nestabilią N-glikozidinę jungtį (34 pav.).
Nukleotidai yra nukleozidų dariniai, fosforilinti vienoje ar keliose ribozės (arba dezoksiribozės) liekanos hidroksilo grupėse. Taigi adenozino monofosfatas (AMP arba adenilatas) yra sudarytas iš adenino, ribozės ir fosfato. 2'-deoksiadenozino monofosfatas (dAMP arba deoksiadenilatas) yra molekulė, sudaryta iš adenino, 2'-dezoksiribozės ir fosfato. Paprastai ribozė yra prijungta prie uracilo, o 2-deoksiribozė - prie timino. Todėl timidilo rūgštis (TMF) susideda iš timino, 2'-dezoksiribozės ir fosfato. Be minėtų nukleotidų formų, buvo rasta ir neįprastos struktūros nukleotidų. Taigi, tRNR molekulėje buvo rastas nukleotidas, kuriame ribozė yra prijungta prie uracilo penktoje padėtyje, tai yra, ne azoto-anglies ryšiu, o anglies-anglies ryšiu. Šio neįprasto papildymo produktas vadinamas pseudouridinu (ψ). tRNR molekulėse taip pat yra dar viena neįprasta nukleotidų struktūra – timinas, sujungtas su ribozės monofosfatu. Šis nukleotidas susidaro jau po tRNR molekulės sintezės metilinant UMP liekaną S-adenozilmetioninu. Pseudouridilo rūgštis (ψMP) taip pat susidaro dėl UMP persitvarkymo po tRNR sintezės.
34 pav. Purino ir pirimidino nukleozidų ir nukleotidų struktūra
Nukleozidų ir nukleotidų nomenklatūra, fizikinės ir cheminės savybės ir funkcijos
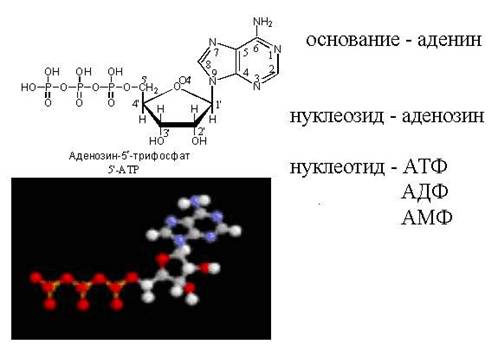
Fosfato grupės padėtis nukleotidų molekulėje nurodoma skaičiumi. Pavyzdžiui, adenozinas su fosfato grupe, prijungta prie 3-osios ribozės anglies atomo, būtų vadinamas 3'-monofosfatu. Pirminis skaičius po skaičiaus dedamas siekiant atskirti anglies skaičių purino arba pirimidino bazėje nuo šio atomo padėties dezoksiribozės liekanoje. Skaičiuojant bazės anglies atomus, pradas nededamas. Nukleotidas 2'-deoksiadenozinas su fosfato likučiu cukraus molekulės anglies atome-5 vadinamas 2'-deoksiadenozin-5'-monofosfatu. Nukleozidai, kurių sudėtyje yra adenino, guanino, citozino, timino ir uracilo, paprastai žymimi atitinkamai A, G, C, T ir Y raidėmis. Raidės d (arba d) buvimas prieš santrumpą rodo, kad nukleozido angliavandenių komponentas yra 2′-deoksiribozė. Guanozinas, turintis 2′-dezoksiribozės, gali būti pažymėtas dG (deoksiguanozinu), o atitinkamas monofosfatas su fosfato grupe, prijungtas prie trečiojo dezoksiribozės anglies atomo, gali būti pažymėtas dG-3′-MF. Paprastai, kai fosfatas yra prijungtas prie ribozės arba dezoksiribozės anglies-5, simbolis 5′ praleidžiamas. Pavyzdžiui, guanozino 5′-monofosfatas paprastai vadinamas GMP, o 2′-deoksiguanozino 5′-monofosfatas sutrumpintas kaip dGMP. Jei prie nukleozido angliavandenių liekanos yra prijungtos 2 arba 3 fosforo rūgšties liekanos, vartojamos santrumpos DF (difosfatas) ir TF (trifosfatas). Taigi, adenozinas + trifosfatas su trimis fosfatų grupėmis angliavandenių 5′ padėtyje bus pažymėtas ATP. Kadangi nukleotidų molekulėse esantys fosfatai yra fosforo anhidridų pavidalu, ty mažos entropijos būsenoje, jie vadinami makroergais (turinčiais daug potencialios energijos). Hidrolizuojant 1 molį ATP į ADP, išsiskiria 7,3 kcal potencialios energijos.
35 pav. cAMP struktūra
Fizikinės ir cheminės nukleotidų savybės
Kadangi nukleotiduose yra azoto bazių, tokios savybės kaip tautomerija ir gebėjimas absorbuotis ultravioletinėje spektro dalyje taip pat būdingos nukleotidams, o azoto bazių ir nukleotidų, turinčių šias bazes, absorbcijos spektrai yra panašūs. Dėl cukraus ir fosforo rūgšties likučių jie yra labiau hidrofiliniai nei azoto bazės. Visi nukleotidai yra rūgštys, nes juose yra fosforo rūgšties likučių.
Natūralių nukleotidų funkcijos
Nukleotidai yra nukleorūgščių (RNR, DNR) monomerai. Į DNR sudėtį įeina dezoksiribonukleotido fosfatai – adenino, timino, guanino ir citozino dariniai. Be to, kai kurios guanino ir citozino molekulės DNR yra metilintos, tai yra, jose yra metilo grupė. Kaip pagrindiniai monomerai, RNR yra ribonukleotidų fosfatų – adenino, uracilo, guanino ir citozino darinių. RNR taip pat yra nukleotidų, kuriuose yra įvairių nedidelių azoto bazių, tokių kaip ksantinas, hipoksantinas, dihidrouridinas ir kt.
Nukleotidai yra kofermentų (NAD, NADP, FAD, kofermento A, metionino-adenozino) monomerai. Kaip kofermentų dalis, jie dalyvauja fermentinėse reakcijose. Ši funkcija bus išsamiau aptarta toliau.
Energija (ATP). ATP atlieka pagrindinio tarpląstelinio laisvosios energijos nešėjo funkciją. Gausiausio laisvojo nukleotido ATP koncentracija žinduolių ląstelėse yra apie 1 mmol/l.
Signalas (cGMP, cAMP)(35 pav.). Ciklinis AMP (3-, 5-adenozino monofosfatas, cAMP), įvairių tarpląstelinių signalų tarpininkas gyvūnų ląstelėse, susidaro iš ATP dėl reakcijos, katalizuojamos adenilato ciklazės. Adenilato ciklazės aktyvumą reguliuoja kompleksas sąveikų, kurių daugelis prasideda per hormonų receptorius. Viduląstelinė cAMP koncentracija (apie 1 µmol/l) yra 3 eilėmis mažesnė už ATP koncentraciją. Ciklinis cGMP (3-, 5-guanozino monofosfatas, cGMP) tarnauja kaip tarpląstelinis tarpląstelinių signalų laidininkas. Kai kuriais atvejais cGMP veikia kaip cAMP antagonistas. cGMP susidaro iš GTP, veikiant guanilatciklazei – fermentui, turinčiam daug bendro su adenilato ciklaze. Guanilatciklazę, kaip ir adenilato ciklazę, reguliuoja įvairūs efektoriai, įskaitant hormonus. Kaip ir cAMP, cGMP yra hidrolizuojamas fosfodiesterazės iki atitinkamo 5'-monofosfato.
Reguliavimo (GTP). Baltymų grupės (G-baltymų), kurios daugiausia atlieka reguliavimo funkciją, aktyvumas priklauso nuo to, kurį nukleotidą jie jungiasi. Neaktyvioje formoje šie baltymai suriša BVP; kai baltymas aktyvuojamas, BVP pakeičiamas GTP. Vykdydamas savo funkciją baltymas hidrolizuoja GTP į BVP ir fosfatą, išsiskirianti energija išleidžiama baltymo funkcionavimui.
Aktyvinimas lipidų ir monosacharidų metabolizmo metu (UTP, STP). Uracilo nukleotidų dariniai dalyvauja kaip aktyvuojantys agentai heksozės metabolizme ir angliavandenių polimerizacijos reakcijose, ypač krakmolo ir glikoproteinų bei proteoglikanų fragmentų oligosacharidų biosintezėje. Šių reakcijų substratai yra uridino difosfato cukrus. Pavyzdžiui, uridino difosfato gliukozė yra glikogeno pirmtakas. Be to, gliukozė virsta galaktoze, gliukurono rūgštimi ar kitais monosacharidų dariniais kaip konjugatas su UDP. CTP reikalingas tam tikrų fosfogliceridų biosintezei gyvūnų audiniuose. Reakcijos, susijusios su keramidu ir CDP-cholinu, sukelia sfingomielino ir kitų pakeistų sfingozinų susidarymą.
Dalyvavimas dezaktyvuojant įvairius alkoholius ir fenolius(UDP-gliukurono rūgštis). Uridino difosfato gliukurono rūgštis – veikia kaip „aktyvus“ gliukuronidas konjugacijos reakcijose, pavyzdžiui, susidarant bilirubino gliukuronidui.
Nukleotidai kofermentuose
Kofermentai yra mažos molekulinės masės junginiai, susiję su fermentais (žr. skyrių „Fermentai“), tiesiogiai dalyvaujančiais biocheminėje reakcijoje, kitaip tariant, tai dar vienas substratas, kuris nepatenka į aplinką.
Kofermentai skirstomi į dvi grupes:
protonų ir elektronų nešėjai, šie kofermentai dalyvauja redokso reakcijose;
visų kitų grupių nešėjai, išskyrus protonus ir elektronus, šie kofermentai dalyvauja transferazės reakcijose.
Išsamiau apie šių reakcijų mechanizmus galima susipažinti skyriuje „Fermentai“.
Kai kurių kofermentų sudėtyje yra nukleotidų. Jie taip pat skirstomi į tas pačias dvi grupes.
kofermentai, pernešantys protonus ir elektronus
Šie kofermentai dalyvauja redokso reakcijose, kur adenozinas atlieka tik struktūrinę funkciją, į reakciją patenka kitų tipų bazių turintys nukleotidai, išskiriami du tokių kofermentų tipai: nikotino ir flavino. Jie skiriasi ne tik aktyvia grupe, bet ir jų vykdomų reakcijų tipu.
Nikotininiai kofermentai
36 pav. Nikotininiai kofermentai. NAD A struktūra, NADP B struktūra, nikotino rūgšties C aktyvumo mechanizmas, nikotino kofermentų D veikimo mechanizmas
Nikotinamido adenino dinukleotidas (NAD+) yra pagrindinis elektronų akceptorius oksiduojant kuro molekules. Reaktyvioji NAD+ dalis yra jo nikotinamido žiedas. Kai substratas oksiduojamas, NAD+ nikotinamido žiedas prideda vandenilio joną ir du elektronus, kurie yra lygiaverčiai hidrido jonui. Sumažinta šio nešiklio forma yra NADH. Šio dehidrogenavimo metu vienas substrato vandenilio atomas tiesiogiai perkeliamas į NAD+, o antrasis – į tirpiklį. Abu substrato prarasti elektronai perkeliami į nikotinamido žiedą. Elektronų donoro vaidmuo daugumoje redukcinės biosintezės (plastinės mainų) procesų; atlieka redukuotą nikotino amido adenino dinukleotido fosfato (NADPH) formą. NADPH nuo NAD skiriasi tuo, kad yra fosfatas, esterintas iki adenozino 2'-hidroksilo grupės. Oksiduota NADPH forma vadinama NADP+. NADPH perneša elektronus taip pat, kaip ir NADH. Tačiau NADPH naudojamas beveik išimtinai redukciniuose biosintezės procesuose, o NADH pirmiausia naudojamas ATP generavimui. Papildoma NADPH fosfatų grupė yra vieta, atsakinga už numatytą molekulės paskirtį, kuri yra fermentų atpažinimas.
Flavino kofermentai
Pirmąjį flavino kofermentą (flavino mononukleotidą FMN) A. Szent-Györgyi išskyrė iš širdies raumens 1932 m., R. G. Warburgas ir V. Christianas tuo pačiu metu gavo pirmąjį flavoproteiną, turintį FMN kaip kofermentą iš mielių. Antrasis svarbiausias flavino kofermentas, flavino adenino dinukleotidas (FAD), buvo išskirtas kaip D-aminorūgšties oksidazės kofaktorius 1938 m. Dėl flavino žiedo redokso transformacijos flavino kofermentai vykdo redokso reakcijas kaip daugelio svarbių fermentų sistemų dalis: oksidazės (ypač D- ir L-aminorūgščių oksidazės, monoaminooksidazė, kuri reguliuoja katecholaminų kiekį organizme). kraujas) ir dehidrogenazės (dažnai dalyvaujant nikotinamido adenino dinukleotidui ir ubichinonams).
37 pav. Flavino kofermentai. FAD A struktūra, nikotino rūgšties aktyvumo B mechanizmas, flavino kofermentų C mechanizmas
Antrasis pagrindinis elektronų nešiklis oksiduojant kuro molekules yra flavino adenino dinukleotidas. Šio nešiklio oksiduotų ir redukuotų formų santrumpos yra atitinkamai FAD ir FADH 2 . Reaktyvioji FAD dalis yra jo izoalloksazino žiedas. FAD, kaip ir NAD +, prideda du elektronus. Tačiau FAD, skirtingai nei NAD +, prideda abu substrato prarastus vandenilio atomus.
Įvadinio segmento pabaiga.
Nukleotidų molekulėje yra cukraus, fosfato ir azoto bazės. Kaip šie paprasti komponentai leidžia nukleotidams susijungti ir sukurti polimerus, tokius kaip DNR ir RNR, taip pat energiją nešančias molekules, tokias kaip ATP?
Nukleotidai: DNR struktūros dalis
Kas yra nukleotidas? Norėdami tai suprasti, turite įsivaizduoti DNR. Patekus į ląstelės branduolį ir išardius chromosomas, galima pamatyti ploną dvigubą grandinę. Priartinus matote, kad kiekviena iš šių gijų sudaryta iš mažų statybinių blokų, vadinamų nukleotidais.
Jei DNR atrodo kaip susuktos kopėčios, kiekvienas statybinis blokas arba nukleotidas apima pusę laiptelio ir tam tikrą vertikalią kopėčių dalį. Kita laiptelio pusė nurodo gretimą DNR grandinę. Nukleotidai taip pat gali egzistuoti patys arba būti kitų svarbių molekulių, išskyrus DNR, dalis. Pavyzdžiui, energijos nešiklis ATP yra nukleotido forma.

Nukleotidų komponentai
Nukleotidas susideda iš tokių komponentų kaip azoto bazė, cukrus ir vienas ar daugiau fosfatų. Verta apsvarstyti kiekvieną iš jų išsamiau:
- azoto bazė. Tai gali būti adeninas, timinas, citozinas, guaninas, uracilas. Jie nėra rūgštys, kiekvienoje iš jų yra keli azoto atomai. Nukleotidai gali poruotis vienas su kitu: citozinas visada poruojasi su guaninu, o adeninas – su timinu DNR arba uracilu RNR.
- Kitas pagrindinis nukleotido komponentas yra cukrus. Yra daug cukraus rūšių, tačiau čia svarbios dvi: ribozė yra cukrus, kurį matysite RNR. Yra ribozės versija, kurioje nėra deguonies atomo ir kuri būtų vadinama cukraus dezoksiriboze. Tai yra cukraus rūšis DNR nukleotiduose. Atminkite, kad DNR yra dezoksiribonukleino rūgštis.
- Paskutinis pagrindinis nukleotido fragmentas yra fosfatas. Fosfatas yra fosforo atomas, sujungtas su keturiais deguonies atomais. Ryšiai tarp fosfatų yra labai energingi ir veikia kaip energijos kaupimo forma. Nutrūkus ryšiui, gautą energiją galima panaudoti darbui atlikti.

Nukleotidų tipai
Kai nukleotidai polimerizuojasi arba susijungia, jie sudaro nukleorūgštį, tokią kaip DNR arba RNR. Kiekvienas nukleotido fosfatas yra prijungtas prie skirtingo cukraus, sudarydamas cukraus-fosfato pagrindą su azotinėmis bazėmis. Nukleozidas yra nukleotido dalis, kurią sudaro tik cukrus ir bazė. Taigi apie nukleotidą galime kalbėti kaip apie nukleozidą ir fosfatus:
- Nukleozido monofosfatas yra nukleotidas, kuriame yra vienas fosfatas.
- Nukleozidų difosfatas yra nukleotidas, kurį sudaro du fosfatai.
- Nukleozidų trifosfatas yra nukleotidas, kuriame yra trys fosfatai. Nukleotidai yra DNR ir RNR statybiniai blokai.
Kokie yra nukleotidų tipai, kokia jų sandara ir kaip vieno nukleotido pokytis gali paveikti organizmo išlikimą?

Nukleotidas yra biologijoje... (apibrėžimas)
Žmogaus DNR sudaro nukleotidai, kurie iš esmės yra DNR subelementinė dimensija, išdėstyta poromis. Šių bazinių porų, dar vadinamų bazinėmis poromis, yra apie 3 milijardai. Koks yra nukleotido apibrėžimas? Kiekvienos spermatozoidų ląstelės ir kiekvieno kiaušinėlio branduolyje yra maždaug šeši milijardai atskirų nukleotidų, kurie yra suskirstyti į kompaktiškas DNR molekules. Dėl to juos lengviau laikyti ir perkelti.
Taigi, kas yra nukleotidai? Jie elgiasi kaip ypatinga kalba, kuris naudojamas jūsų organizmo gaminamų cheminių medžiagų, ypač baltymų, receptams rašyti. Dauguma nukleotidų sekcijų vadinamos nepageidaujama DNR, nes jos nieko nekoduoja. Tačiau yra nedidelė dalis, kuri yra labai svarbi jūsų išlikimui ir padaro jus tuo, kas esate. Tai 2% nukleotidų kodas kiekvienam baltymui, kurį jūsų kūnas gamina ir turi DNR skyriuose, vadinamuose genais. Kiekvienas genas koduoja aminorūgščių grandinę, kuri veda prie specifinio baltymo susidarymo.
Mutacijos, kurios yra ląstelės DNR pokyčiai, apimantys vieną nukleotidą, gali atrodyti nereikšmingi, atsižvelgiant į tai, kad žmogaus genome yra tiek daug nukleotidų, tačiau kai jos atsiranda tam tikruose genuose, jos gali sukelti gyvybei pavojingas ligas. Norint geriau suprasti šį mechanizmą, pirmiausia reikia pažvelgti į kai kuriuos nukleotidų pagrindus.
Nukleotidų struktūra
Nukleotidai yra monomerai (arba Statybiniai blokai) nukleorūgštys ir susideda iš 5 anglies cukraus, fosfatų grupės ir azoto bazės. Kaip jau minėta, cukrus ir bazė kartu sudaro nukleozidą. Fosfato grupės pridėjimas paverčia molekulę nukleotidu. Nukleotidai įvardijami pagal juose esančią azoto bazę ir prie jos prijungtą cukrų (pavyzdžiui, dezoksiribozė DNR nukleotiduose ir ribozė RNR). Kokie nukleotidai yra DNR ir RNR? DNR ir RNR yra aštuoni skirtingi nukleozidai:
- RNR: adenozinas, guanozinas, citidinas, uridinas.
- DNR: deoksiadenozinas, deoksiguanozinas, deoksicitidinas, deoksitimidinas.
Yra ir kitų svarbių nukleotidų, pvz., dalyvaujančių metabolizme (pvz., ATP) ir ląstelių signalizacijoje (pvz., GTP).

Nukleotidų surišimas
Norint sukurti polimerų grandines (arba kelis vienetus), kurios veda į RNR ir DNR susidarymą, nukleotidai yra sujungti vienas su kitu per cukraus fosfato pagrindą, kuris susidaro, kai vieno nukleotido fosfatas yra prijungtas prie kito nukleotido cukraus. Tai įmanoma dėl stiprių kovalentinių ryšių, vadinamų fosfodiesterio ryšiais.
Kadangi DNR yra dvigrandė molekulė, dvi iš šių polimerinių grandinių turi susijungti kaip kopėčios. „Laiptai“ yra sudaryti iš nukleotidų porų, jungiančių dvi kopėčių puses vandeniliniais ryšiais. Kas yra nukleotidas? Tai struktūrinis DNR vienetas, susidedantis iš azoto bazės ir cukraus-fosfato pagrindo, susidedančio iš fosfatų grupės ir cukraus. DNR sudaro daugybė nukleotidų, kuriuose yra ir saugomi organizmo genetiniai kodai.

Nukleino rūgštys
Nukleorūgštys yra biopolimerai, kurie kartu su baltymais žaidžia svarbus vaidmuo visų gyvų organizmų ląstelėse. Šie junginiai yra atsakingi už paveldimos informacijos saugojimą, perdavimą ir įgyvendinimą. Kas yra nukleotidai? Jie yra nukleorūgščių monomerai.
Tarp nukleotido dalių susidaro kovalentinės cheminės jungtys, kurios susidaro dėl kondensacijos reakcijų. Tokios reakcijos yra atvirkštinė hidrolizė. Įdomus faktas yra tai, kad DNR molekulės paprastai yra ne tik ilgesnės už RNR molekules, bet ir apima dvi grandines, kurios yra sujungtos viena su kita vandeniliniais ryšiais, atsirandančiais tarp azoto bazių.
yra sudėtingi monomerai, iš kurių surenkamos heteropolimerų molekulės. DNR ir RNR. Laisvieji nukleotidai dalyvauja gyvybės signalų ir energijos procesuose. DNR nukleotidai ir RNR nukleotidai turi bendrą struktūrinį planą, tačiau skiriasi pentozės cukraus struktūra. DNR nukleotidai naudoja cukraus dezoksiribozę, o RNR nukleotidai naudoja ribozę.
Nukleotido struktūra
Kiekvienas nukleotidas gali būti suskirstytas į 3 dalis:
1. Angliavandenis yra penkių narių pentozės cukrus (ribozė arba dezoksiribozė).
2. Fosforo liekana (fosfatas) – tai fosforo rūgšties likutis.
3. Azoto bazė – tai junginys, kuriame yra daug azoto atomų. Nukleino rūgštyse naudojamos tik 5 azoto bazių rūšys: adeninas, timinas, guaninas, citozinas, uracilas. DNR yra 4 tipai: adeninas, timinas, guaninas, citozinas. RNR taip pat yra 4 tipai: Adeninas, Uracilas, Guaninas, Citozinas Nesunku pastebėti, kad RNR lyginant su DNR timinas yra pakeistas Uracilu.
Bendra struktūrinė pentozės (ribozės arba dezoksiribozės) formulė, kurios molekulės sudaro nukleorūgščių „skeletą“:
Jei X yra pakeistas H (X = H), tada gaunami dezoksiribonukleozidai; jei X pakeičiamas OH (X = OH), tai gaunami ribonukleozidai. Jei vietoj R pakeisime azoto bazę (puriną arba pirimidiną), gausime specifinį nukleotidą.
Svarbu atkreipti dėmesį į tas anglies atomų pozicijas pentozėje, kurios žymimos 3" ir 5". Anglies atomų numeracija prasideda nuo deguonies atomo viršuje ir eina pagal laikrodžio rodyklę. Gaunamas paskutinis anglies atomas (5"), kuris yra už pentozės žiedo ir sudaro, galima sakyti, pentozės "uodegą". Taigi, sudarydamas nukleotidų grandinę, fermentas gali prijungti tik naują nukleotidą. anglies 3 "ir jokiam kitam . Todėl 5 colių nukleotidų grandinės galas niekada negali būti tęsiamas; tik 3 colių galas gali būti pailgintas.



Palyginkite RNR nukleotidą su DNR nukleotidu.
Pabandykite išsiaiškinti, koks tai nukleotidas šiame paveikslėlyje:

ATP – laisvasis nukleotidas
cAMP – „atgalinė“ ATP molekulė

Nukleotidų struktūros diagrama

Atkreipkite dėmesį, kad aktyvuotas nukleotidas, galintis sukurti DNR arba RNR grandinę, turi „trifosfato uodegą“. Būtent su šia „energija prisotinta“ uodega ji gali prisijungti prie jau esančios augančios nukleino rūgšties grandinės. Fosfato uodega yra ant anglies 5, todėl anglies vieta jau yra užimta fosfatų ir yra skirta pritvirtinti. Prie ko pritvirtinti? Tik prie anglies, esančios 3 padėtyje". Prisijungęs šis nukleotidas pats taps kito nukleotido taikiniu. „Priimantioji pusė" suteikia 3 padėtyje esančią anglį, o „atvykstanti pusė" prie jos prilimpa fosfato uodega, esanti 5" padėtyje. Apskritai grandinė auga iš 3" pusės.
DNR nukleotidų grandinės pratęsimas
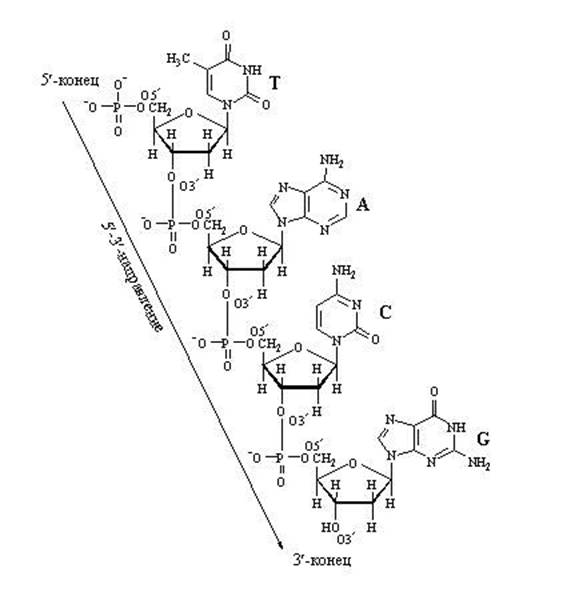
Grandinės augimas dėl „išilginių“ ryšių tarp nukleotidų gali vykti tik viena kryptimi: nuo 5" ⇒ iki 3", nes Naują nukleotidą galima pridėti tik prie 3' grandinės galo, o ne į 5' galą.
Nukleotidų poros, sujungtos "kryžminiais" komplementariais jų azotinių bazių ryšiais

DNR dvigubos spiralės pjūvis
Raskite dviejų DNR grandžių antilygiagretumo požymius.
Raskite nukleotidų poras su dvigubomis ir trigubomis komplementariomis jungtimis.
Nukleotidai
Pažvelkime atidžiau į nukleotidus. Yra žinoma, kad nukleotidai vadinami adeninu, guaninu, timinu, citozinu ir uracilu - azoto bazėmis, jie parodyti paveikslėlyje žemiau.
Nukleotidai yra nukleorūgščių monomerai. Nukleorūgštys eukariotinėse ląstelėse yra branduolyje. Visi gyvi organizmai jas turi (neturintys branduolio dar turi nukleino rūgščių – jos bakterijose yra ląstelės centre ir formuoja nukleoidus). Monomerai, iš kurių susidaro nukleorūgštys, susideda iš azoto bazės, cukraus liekanos (dezoksiribozės arba ribozės) ir fosfato. Cukrus kartu su azotine baze vadinami nukleozidais (adenozinas, guanozinas, timidinas, citidinas). Jei prie jų yra prijungtos 1-, 2- arba 3-fosforo liekanos, visa ši struktūra vadinama atitinkamai nukleotido monofosfatu, difosfatu arba trifosfatu arba nukleotidu (adeninu, guaninu, timinu, citozinu).
Taip ATP modelis atrodo erdvėje. Azoto bazė, kuri yra DNR dalis, yra padalinta į dvi grupes - pirimidiną ir puriną. DNR yra adenino, timino, citozino ir guanino; RNR yra uracilas, o ne timinas. Kaip žinote, DNR yra didelis archyvas, kuriame saugoma informacija, o RNR yra molekulė, kuri perduoda informaciją iš branduolio į citoplazmą baltymų sintezei. Funkcijų skirtumai yra susiję su struktūros skirtumais. RNR yra chemiškai aktyvesnė dėl to, kad jos cukrus - ribozė - turi hidroksilo grupę, o deoksiribozėje nėra deguonies. Dėl deguonies trūkumo DNR yra inertiškesnė, o tai svarbu jos informacijos saugojimo funkcijai, kad ji nepatektų į jokias reakcijas.
Nukleotidai gali sąveikauti vienas su kitu, o du fosforai yra „išmesti“, o tarp gretimų nukleotidų susidaro ryšys. Furanozės molekulėje anglies molekulės yra sunumeruotos. Azoto bazė yra susijusi su pirmuoju. Kai susidaro nukleotidų grandinė, ryšys yra tarp penktosios vienos ir trečiosios kitos fosforo rūgšties anglies. Todėl nukleorūgščių grandinėje išskiriami skirtingi nelygūs galai, kurių atžvilgiu molekulė nėra simetriška.
Komplementarios vienos grandinės nukleino rūgšties molekulės gali sudaryti dvigrandę struktūrą. Šioje spiralėje adeninas poruojasi su timinu, o guaninas – su citozinu. Yra teiginys, kad nukleotidai dera kaip fragmentai dužęs stiklas todėl jie sudaro poras. Tačiau šis teiginys yra klaidingas. Nukleotidai gali sudaryti poras bet kokiu būdu. Vienintelė priežastis, dėl kurios jie susijungia taip ir ne kas kita, yra ta, kad kampas tarp „uodegų“, einančių į cukrų, sutampa tik šiose porose, be to, sutampa jų dydžiai. Jokia kita pora nesudaro tokios konfigūracijos. Ir kadangi jie sutampa, jie gali būti sujungti vienas su kitu per cukraus ir fosfato stuburą. Dvigubos spiralės struktūrą 1953 m. atrado Jamesas Watsonas ir Francisas Crickas.
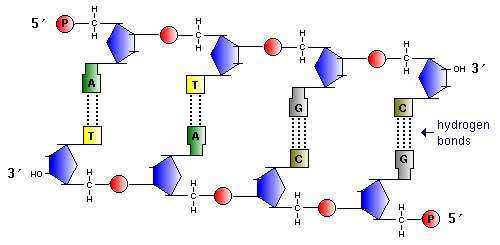
Sujungus vienas su kitu, priešais vienos gijos 5' galą yra kitos gijos 3' galas. Tai yra, sruogos eina priešingomis kryptimis – sakoma, kad DNR grandinės yra antilygiagrečios.
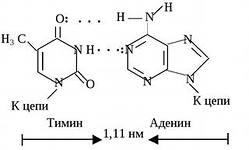
Paveikslėlyje parodytas DNR modelis, matyti, kad adeninas su timinu yra sujungtas dviem vandeniliniais ryšiais, o guaninas su citozinu – trigubu vandenilio ryšiu. Jei DNR molekulė kaitinama, aišku, kad du ryšius nutraukti lengviau nei tris, o tai būtina DNR savybėms.
Dėl cukraus ir fosfato stuburo ir nukleotidų erdvinio išsidėstymo, kai nukleotidai sukraunami vienas ant kito ir „persiūti“ per cukraus ir fosfato stuburą, grandinė pradeda vyniotis, taip suformuodama garsiąją dvigubą spiralę.
Paveiksluose pavaizduoti rutuliniai DNR modeliai, kur kiekvienas atomas pažymėtas rutuliuku. Spiralės viduje yra grioveliai: maži ir dideli. Per šiuos griovelius baltymai sąveikauja su DNR ir atpažįsta ten esančią nukleotidų seką.
Kai DNR kaitinama, vandeniliniai ryšiai nutrūksta ir dvigubos spiralės gijos išsiskleidžia. Kaitinimo procesas vadinamas DNR tirpimu, o ryšiai tarp A-T ir G-C porų sunaikinami. Kuo daugiau DNR garas A-T, kuo mažiau tvirtai sruogos sujungtos viena su kita, tuo lengviau ištirpsta DNR. Perėjimas nuo dvigrandės DNR į viengrandę DNR matuojamas spektrofotometrais šviesos sugerties būdu esant 260 nm. DNR lydymosi temperatūra priklauso nuo A-T/G-C sudėties ir molekulės fragmento dydžio. Akivaizdu, kad jei fragmentas susideda iš kelių dešimčių nukleotidų, tada jis ištirpsta daug lengviau nei ilgesni fragmentai.
Žmogaus haploidiniame genome, tai yra viename chromosomų rinkinyje, yra 3 milijardai nukleotidų porų, jų ilgis yra 1,7 m, o ląstelė yra daug mažesnė, kaip galite spėti. Kad DNR galėtų joje tilpti, ji yra tvirtai susilanksčiusi, o eukariotų ląstelėje susilankstyti padeda baltymai – histonai. Histonai yra teigiamai įkrauti, o kadangi DNR yra neigiamai įkrauta, histonai turi afinitetą DNR. Histonu supakuota DNR yra granulių, vadinamų nukleosomomis, pavidalu. 200 nukleotidų porų patenka į vieną nukleozomą, 146 poros yra suvyniotos aplink histonus, o likusios 54 poros kabo linkerinės (surišančios nukleozomų) DNR pavidalu. Tai pirmasis DNR sutankinimo lygis. Chromosomose DNR sulankstoma dar kelis kartus, kad susidarytų kompaktiškos struktūros.
Be DNR, RNR taip pat priklauso nukleino rūgštims. Ląstelėje yra įvairių tipų RNR: ribosominės, šabloninės, transportinės. Yra ir kitų RNR tipų, apie kuriuos kalbėsime vėliau. RNR sintetinama kaip vienagrandė molekulė, tačiau kai kurios jos dalys yra dvigrandžių spiralių dalis. Kalbant apie RNR, taip pat kalbama apie pirminę struktūrą (nukleotidų sekas) ir antrinę struktūrą (dvigrandžių skyrių susidarymą).
Lipidai
Lipidai yra sudaryti iš riebalų rūgščių su ilgomis angliavandenilių grandinėmis. Riebalų rūgštys yra hidrofobinės, tai reiškia, kad jos netirpsta vandenyje.
Lipidai yra riebalų rūgščių junginiai su gliceroliu (esteriais). Pavyzdžiui, paveikslėlyje pavaizduotas lecitinas.
Svarbų vaidmenį ląstelėje atlieka lipidai, kuriuose prie glicerolio prisijungia fosforo rūgšties liekana ir 2 riebalų rūgštys. Jie vadinami fosfolipidais. Fosfolipidų molekulės turi polinę (ty hidrofilinę, labai tirpią) grupę viename molekulės gale ir ilgą hidrofobinę uodegą. Fosfolipidai apima fosfatidilcholiną.
Vandeniniame tirpale fosfolipidai sudaro miceles, kuriose molekulės savo polines „galvas“ pasuka į išorę, vandens link, o hidrofobinės „uodegos“ yra micelių viduje, paslėptos nuo vandens. Ląstelės membrana taip pat yra lipidinė su polinėmis „galvomis“, kurios yra nukreiptos į išorę abiejose membranos pusėse, o hidrofobinės „uodegos“ yra lipidų dvigubo sluoksnio viduje.
Bibliografija
Rengiant šį darbą buvo naudojama medžiaga iš svetainės http://bio.fizteh.ru.
Iki 1944 m. O. Avery ir jo kolegos K. McLeod ir M. McCarthy atrado transformuojantį pneumokokų DNR aktyvumą. Šie autoriai tęsė Griffith darbą, kuris aprašė transformacijos (paveldimų savybių perkėlimo) reiškinį bakterijose. O. Avery, K. McLeod, M. McCarthy parodė, kad pašalinus baltymus, polisacharidus ir RNR, bakterijų transformacija nesutrikdoma, o kai indukuojančią medžiagą veikia fermentas dezoksiribonukleazė, transformuojantis aktyvumas išnyksta.
Šiuose eksperimentuose pirmą kartą buvo įrodytas genetinis DNR molekulės vaidmuo. 1952 metais A. Hershey ir M. Chase'as patvirtino genetinį DNR molekulės vaidmenį eksperimentuose su T2 bakteriofagu. Pažymėję jos baltymą radioaktyvia siera, o DNR – radioaktyviuoju fosforu, jie šiuo bakteriniu virusu užkrėtė E. coli. Fago palikuoniuose buvo aptiktas didelis kiekis radioaktyvaus fosforo ir tik pėdsakai S. Iš to seka, kad į bakteriją prasiskverbia ne fago baltymas, o DNR, o po replikacijos perkeliami į fago palikuonis.
DNR nukleotido sandara. Nukleotidų tipai.
Nukleotidas DNR sudaryta iš
Azoto bazė (4 tipai DNR: adeninas, timinas, citozinas, guaninas)
Monocukraus dezoksiribozė
Fosforo rūgštis
nukleotidų molekulė susideda iš trijų dalių – penkių angliavandenių cukraus, azoto bazės ir fosforo rūgšties.
Cukrus įtrauktas į nukleotidų sudėtis, yra penki anglies atomai, tai yra, tai yra pentozė. Priklausomai nuo nukleotide esančios pentozės tipo, yra dviejų tipų nukleorūgštys – ribonukleorūgštys (RNR), kuriose yra ribozės, ir dezoksiribonukleorūgštys (DNR), kuriose yra dezoksiribozės. Dezoksiribozėje OH grupė 2-ajame anglies atome pakeičiama H atomu, tai yra, joje yra vienu deguonies atomu mažiau nei ribozėje.
Abejuose nukleino rūgščių rūšys yra keturių pagrindų skirtingi tipai: du iš jų priklauso purinų klasei, o du – pirimidinų klasei. Žiede esantis azotas suteikia šiems junginiams pagrindinį pobūdį. Purinai apima adeniną (A) ir guaniną (G), o pirimidinai – citoziną (C) ir timiną (T) arba uracilą (U) (atitinkamai DNR arba RNR). Timinas chemiškai yra labai artimas uracilui (tai 5-metiluracilas, tai yra uracilas, kuriame metilo grupė yra 5-ame anglies atome). Purino molekulė turi du žiedus, o pirimidino molekulė turi vieną.
Nukleotidai yra sujungti stipriu kovalentiniu ryšiu per vieno nukleotido cukrų ir kito fosforo rūgštį. Paaiškėja polinukleotidinė grandinė. Viename gale yra laisva fosforo rūgštis (5' galas), kitame - laisvas cukrus (3' galas). (DNR polimerazė gali pridėti naujų nukleotidų tik prie 3' galo.)
Dvi polinukleotidų grandinės yra sujungtos viena su kita silpnais vandeniliniais ryšiais tarp azoto bazių. Yra 2 taisyklės:
komplementarumo principas: timinas visada priešingas adeninui, guaninas visada priešingas citozinui (jie atitinka viena kitą vandenilinių jungčių forma ir skaičiumi – tarp A ir G yra du ryšiai, o tarp C ir G – 3).
antiparalelizmo principas: kai viena polinukleotidinė grandinė turi 5' galą, kita - 3' galą ir atvirkščiai.
Paaiškėja dviguba grandinė DNR.
Ji susisuka į dviguba spiralė, vienas spiralės posūkis yra 3,4 nm ilgio, jame yra 10 nukleotidų porų. Azoto bazės (genetinės informacijos saugotojai) yra spiralės viduje, saugomos.
DNR molekulės struktūrinė organizacija. J. Watson ir F. Crick modelis
1950 metais anglų fizikas M. Wilkinsas gavo kristalinių DNR skaidulų rentgeno nuotrauką. Ji parodė, kad DNR molekulė turi tam tikrą struktūrą, kurios dekodavimas padėtų suprasti DNR funkcionavimo mechanizmą. Rentgeno spindulių piešiniai, gauti ne ant kristalinių DNR skaidulų, o ant mažiau tvarkingų agregatų, kurie susidaro esant didesnei drėgmei, leido M. Wilkinso kolegei Rosalind Franklin pamatyti aiškų kryžminį raštą – dvigubos spiralės identifikavimo ženklą. Taip pat tapo žinoma, kad nukleotidai yra vienas nuo kito 0,34 nm atstumu, o viename spiralės posūkyje jų yra 10. DNR molekulės skersmuo yra apie 2 nm. Tačiau iš rentgeno spindulių difrakcijos duomenų nebuvo aišku, kaip grandinės laikomos kartu DNR molekulėse.
Vaizdas tapo visiškai aiškus 1953 m., kai amerikiečių biochemikas J. Watsonas ir anglų fizikas F. Crickas, tirdami DNR molekulės struktūrą, priėjo prie išvados, kad cukraus-fosfato stuburas yra DNR molekulės periferijoje. o purino ir pirimidino bazės yra viduryje. Be to, pastarosios yra orientuotos taip, kad tarp bazių iš priešingų grandinių gali susidaryti vandeniliniai ryšiai. Iš jų sukurto modelio paaiškėjo, kad bet kuris purinas vienoje grandinėje visada yra vandeniliu prijungtas prie vienos iš pirimidinų kitoje grandinėje. Tokios poros yra vienodo dydžio per visą molekulės ilgį. Lygiai taip pat svarbu, kad adeninas gali poruotis tik su timinu, o guaninas – tik su citozinu. Šiuo atveju tarp adenino ir timino susidaro dvi vandenilinės jungtys, o tarp guanino ir citozino – trys.
DNR savybės ir funkcijos.
Paveldimos informacijos saugojimas (genetinis kodas – būdas įrašyti genetinę informaciją apie aminorūgščių seką baltyme naudojant nukleotidus (Gamow)
Perkėlimas (pakartoti / padvigubinti)
Diegimas (transkripcija)
DNR autoreprodukcija. Replikonas ir jo veikimas.
Nukleino rūgšties molekulių savaiminio dauginimosi procesas, lydimas tikslių genetinės informacijos kopijų perdavimo (iš ląstelės į ląstelę) paveldėjimo būdu; atliekamas dalyvaujant specifinių fermentų rinkiniui (helikazei, kuri kontroliuoja DNR molekulės išsivyniojimą, DNR polimerazę, DNR ligazę), praeina per pusiau konservatyvų tipą, susidarant replikacijos šakutei; vienoje iš grandinių papildomos grandinės sintezė yra nenutrūkstama, o kitoje - dėl Dkazaki fragmentų susidarymo. Didelio tikslumo procesas, kurio klaidų lygis neviršija 10 -9; eukariotuose jis gali atsirasti vienu metu keliuose vienos DNR molekulės taškuose; greitis eukariotuose yra apie 100, o bakterijų – apie 1000 nukleotidų per sekundę.
Replikonas yra genomo srities replikacijos proceso vienetas, kurį kontroliuoja vienas replikacijos pradžios (pradžios) taškas. Terminą F. Jacobas ir S. Brenneris pasiūlė 1963 m. Prokariotų genomas paprastai yra vienas replikonas. Nuo inicijavimo taško replikacija vyksta abiem kryptimis, kai kuriais atvejais nevienodu greičiu. Eukariotų genomas susideda iš daugybės (dažnai iki kelių dešimčių tūkstančių) replikonų.
Genetinis kodas, jo savybės.
Genetinis kodas yra būdas įrašyti genetinę informaciją apie aminorūgščių seką baltyme naudojant nukleotidus. Geno atradimas Kodas priklauso Georgijui Gamow. 1954 m
Trigubai- reikšmingas kodo vienetas yra trijų nukleotidų derinys (tripletas arba kodonas).
Tęstinumas- tarp trynukų nėra skyrybos ženklų, tai yra, informacija skaitoma nuolat.
nesutampa- tas pats nukleotidas vienu metu negali būti dviejų ar daugiau tripletų dalis (nepastebėta kai kuriems persidengiantiems virusų, mitochondrijų ir bakterijų genams, koduojantiems kelis kadrų poslinkio baltymus).
Vienareikšmiškumas (specifiškumas)- tam tikras kodonas atitinka tik vieną aminorūgštį (tačiau UGA kodonas in Euplotes crassus koduoja dvi aminorūgštis - cisteiną ir selenocisteiną)
Degeneracija (redundancija) Tą pačią aminorūgštį gali atitikti keli kodonai.
Universalumas- genetinis kodas vienodai veikia įvairaus sudėtingumo organizmuose – nuo virusų iki žmonių (tuo pagrįsti genų inžinerijos metodai; yra nemažai išimčių, pateiktų lentelėje „Standartinio genetinio kodo variantai“). “ skyrių žemiau).
Triukšmo atsparumas- nukleotidų pakaitų mutacijos, kurios nelemia koduojamos aminorūgšties klasės pasikeitimo, vadinamos konservatyvus; vadinamos nukleotidų pakeitimo mutacijos, dėl kurių pasikeičia koduojamos aminorūgšties klasė radikalus.
Geno samprata. Genų savybės.
Gene- gyvų organizmų paveldimumo struktūrinis ir funkcinis vienetas. Genas yra DNR seka, nurodanti tam tikro polipeptido arba funkcinės RNR seką. Genai lemia paveldimus organizmų požymius, kurie dauginimosi metu perduodami iš tėvų palikuonims. Tuo pačiu metu kai kurios organelės (mitochondrijos, plastidės) turi savo DNR, kuri nėra įtraukta į organizmo genomą, o tai lemia jų savybes.
(Šį terminą 1909 m. įvedė danų botanikas Wilhelmas Johansenas)
diskretiškumas – genų nesuderinamumas;
stabilumas – gebėjimas išlaikyti konstrukciją;
labilumas – gebėjimas pakartotinai mutuoti;
daugybinis alelizmas – populiacijoje egzistuoja daug genų įvairiomis molekulinėmis formomis;
alelizmas – diploidinių organizmų genotipe tik dvi geno formos;
specifiškumas – kiekvienas genas koduoja savo požymį;
pleiotropija – daugybinis geno poveikis;
ekspresyvumas – geno raiškos požymyje laipsnis;
penetrancija – geno pasireiškimo fenotipe dažnis;
amplifikacija – geno kopijų skaičiaus padidėjimas.
Eukariotų genomo organizavimo ypatumai.
eukariotų genomas:
daug genų
daugiau DNR,
chromosomos turi labai sudėtingą genų veiklos valdymo laike ir erdvėje sistemą, susijusią su ląstelių ir audinių diferenciacija organizmo ontogenezėje.
DNR kiekis chromosomose yra didelis ir didėja, kai organizmai tampa sudėtingesni. Eukariotai taip pat turi genų perteklius. Taigi žmogaus genome yra nukleotidų porų, kurių pakanka daugiau nei 2 milijonams struktūrinių genų susidaryti, o žmogaus, 2000 m. duomenimis, 31 tūkstantis visų genų.
Daugiau nei pusė eukariotų genomo haploidinio rinkinio yra unikalūs genai, pristatyta tik vieną kartą. Žmogaus organizme tokių unikalių genų yra 64 proc., blauzdoje – 55 proc., Drosofiloje – 70 proc.
Nukleotidų sekų klasės eukariotų DNR, jų charakteristikos, savybės ir biologinė reikšmė.
Nukleotidų sekos eukariotų genome
60-ųjų pabaigoje amerikiečių mokslininkų R. Britteno, E. Davidsono ir kitų darbai atrado esminį eukariotų genomo molekulinės struktūros bruožą – įvairaus pakartojamumo laipsnio nukleotidų sekas. Šis atradimas buvo atliktas naudojant molekulinį biologinį metodą, tiriant denatūruotos DNR denatūracijos kinetiką. Eukariotų genome išskiriamos šios frakcijos.
1. Unikalūs, t.y. sekos, pateiktos vienu egzemplioriumi arba keliais egzemplioriais. Paprastai tai yra cistronai - struktūriniai genai, koduojantys baltymus.
2. Žemo dažnio pakartojimai – sekos kartojamos dešimtis kartų.
3. Vidutinio, arba vidutinio dažnio, pakartojimai – šimtus ir tūkstančius kartų kartojamos sekos. Tai apima rRNR genus (žmonėms, 200 viename haploidiniame rinkinyje, pelėms, 100, katėms, 1000, žuvims ir žydintiems augalams, tūkstančius), tRNR, ribosomų baltymų ir histono baltymų genus.
4. Aukšto dažnio pasikartojimai, kurių skaičius siekia 10 milijonų (vienam genomui). Tai trumpos (~ 10 bp) nekoduojančios sekos, kurios yra pericentromerinio heterochromatino dalis.
Eukariotų genomo organizavimo lygiai.
Cheminė ir struktūrinė chromosomų sudėtis.
Molekulinės biologijos tyrimai leido susidaryti vaizdą ne tik apie cheminę chromosomų struktūrą, bet ir apie jų supramolekulinę organizaciją bei funkcionavimo ypatybes. Dabar žinoma, kad chromosomos yra nukleoproteinų dariniai, susidedantys iš DNR ir baltymų. Be to, chromosomose yra tam tikras kiekis RNR, susidariusios transkripcijos metu, ir Ca + ir Mg + jonų. Kiekvienoje chromatidėje ir laiko intervale tarpfazės ir chromosomos anafazės-S periodas turi vieną DNR molekulę, kuri lemia visas chromosomos funkcijas, susijusias su paveldimos informacijos saugojimu, jos perdavimu ir įgyvendinimu. Chromosomose esanti DNR molekulė yra glaudžiai susijusi su dviem baltymų klasėmis – histonais (baziniais baltymais) ir nehistonais (rūgštiniais baltymais). Histonai yra maži baltymai, turintys daug įkrautų aminorūgščių (lizino ir arginino). Grynasis teigiamas krūvis leidžia histonams prisijungti prie DNR, nepaisant nukleotidų sudėties. Jie daugiausia priklauso struktūrinei funkcijai. Tai labai stabilūs baltymai, kurių molekulės gali išlikti visą ląstelės gyvenimą. Eukariotų ląstelėje yra 5 histonų tipai, kurie skirstomi į dvi pagrindines grupes: pirmoji grupė (jie žymimi H2A, H2B, H3, H4) yra atsakinga už specifinių dezoksiribonukleoproteinų kompleksų – nukleozomų – susidarymą. Antroji histonų grupė (HI) yra tarp nukleozomų ir fiksuoja nukleozominės grandinės sulankstymą daugiau aukštas lygis struktūrinė organizacija (supernukleozomų siūlas). Tarp histono baltymų, be struktūrinių, yra tokių, kurie gali apriboti DNR prieinamumą DNR surišantiems reguliuojantiems baltymams ir taip dalyvauti reguliuojant genų aktyvumą. Ne histoniniai baltymai yra labai įvairūs. Jų frakcijų skaičius viršija 100. Jų chromosomose yra mažesniais kiekiais, palyginti su histonais, ir daugiausia atlieka reguliavimo funkciją. Jie dalyvauja reguliuojant genų transkripcijos aktyvumą, užtikrinant DNR replikaciją ir taisymą. Daugumos nehistoninių chromatino baltymų ląstelėse yra nedidelis kiekis (nežymus) – tai reguliuojantys baltymai, atpažįstantys specifines DNR sekas ir prie jų prisijungiantys. Jie dalyvauja daugelyje genetinių procesų, tačiau apie juos mažai žinoma. Kiekybiškai vyrauja nehistoniniai baltymai (pagrindiniai), labai mobilūs, santykinai mažo dydžio, turintys didelį elektros krūvį – jie visada jungiasi su nukleozomomis, kuriose yra aktyvių genų. Be to, daugelis fermentų yra įtraukti į nehistoninių baltymų grupę.
Paveldimos medžiagos pakavimo lygiai eukariotuose.
Taigi, DNR pakavimo lygiai yra tokie:
1) Nukleosominė (2,5 dvigrandės DNR posūkio aplink aštuonias histono baltymų molekules).
2) Supernukleosominė – chromatino spiralė (chromonema).
3) Chromatid – spiralizuota chromonema.
4) Chromosoma – ketvirtasis DNR spermalizacijos laipsnis.
Tarpfaziniame branduolyje chromosomos yra dekondensuotos ir jas vaizduoja chromatinas. Despiralizuota sritis, kurioje yra genų, vadinama euchromatinu (laisvu, pluoštiniu chromatinu). Tai būtina sąlyga transkripcijai. Poilsio metu tarp padalijimų tam tikros chromosomų dalys ir visos chromosomos išlieka kompaktiškos.
Šios spiralizuotos, stipriai nudažytos sritys vadinamos heterochromatinu. Jie neaktyvūs transkripcijai. Yra fakultatyvinis ir konstitucinis heterochromatinas.
Fakultatyvinis heterochromatinas yra informatyvus, nes yra genų ir gali patekti į euchromatiną. Iš dviejų homologinių chromosomų viena gali būti heterochromatinė. Konstitucinis heterochromatinas visada yra heterochromatinis, neinformatyvus (neturi genų), todėl visada yra neaktyvus transkripcijos atžvilgiu.
Chromosominė DNR susideda iš daugiau nei 10 8 bazinių porų, iš kurių susidaro informaciniai blokai – tiesiškai išsidėstę genai. Jie sudaro iki 25% DNR. Genas yra funkcinis DNR vienetas, kuriame yra informacijos apie polipeptidų arba visos RNR sintezę. Tarp genų yra tarpikliai – neinformatyvūs skirtingo ilgio DNR segmentai. Genų perteklius atstovauja daugybei - 10 4 identiškų kopijų. Pavyzdys yra t-RNR, rRNR, histonų genai. DNR yra tų pačių nukleotidų sekos. Tai gali būti vidutiniškai pasikartojančios ir labai pasikartojančios sekos. Vidutiniškai pasikartojančios sekos siekia 300 bazių porų su pasikartojimais 10 2 - 10 4 ir dažniausiai atstovauja tarpiklius, perteklinius genus.
Labai pasikartojančios sekos (10 5 - 10 6) sudaro konstitucinį heterochromatiną. Apie 75% viso chromatino nedalyvauja transkripcijoje, jis patenka į labai pasikartojančias sekas ir netranskribuotus tarpiklius.
Metafazės chromosomos morfologiniai ypatumai.
Mitozinis chromatino superkompaktavimas leidžia ištirti chromosomų išvaizdą naudojant šviesos mikroskopiją. Pirmoje mitozės pusėje jie susideda iš dviejų chromatidžių, sujungtų viena su kita pirminio susiaurėjimo srityje ( centromerai arba kinetochore) specialiai organizuota chromosomos dalis, bendra abiem seserinėms chromatidėms. Antroje mitozės pusėje chromatidės atsiskiria viena nuo kitos. Jie sudaro atskiras sruogas. dukterinės chromosomos, paskirstytas tarp dukterinių ląstelių.
Priklausomai nuo centromeros vietos ir abiejose jo pusėse išsidėsčiusių rankų ilgio, išskiriamos kelios chromosomų formos: lygiarankės arba metacentrinės (su centromera viduryje), nelygiašakės arba submetacentrinės (su centromeras pasislinkęs į vieną iš galų), lazdelės formos arba akrocentrinis (su centromeru, esančiu beveik chromosomos gale), ir taškuotas – labai mažas, kurio formą sunku nustatyti (3.52 pav.). Taikant įprastinius chromosomų dažymo metodus, jos skiriasi forma ir santykiniu dydžiu. Taikant diferencinio dažymo metodus, nustatomas nevienodas fluorescencijos arba dažų pasiskirstymas per visą chromosomos ilgį, griežtai specifinis kiekvienai atskirai chromosomai ir jos homologui (3.53 pav.).
Taigi kiekviena chromosoma yra individuali ne tik joje esančių genų rinkinio, bet ir morfologijos bei diferencinio dažymo pobūdžio požiūriu.
Eu- ir heterochromatinas, jų biologinė reikšmė.
Kai kurios chromosomos ląstelių dalijimosi metu atrodo kondensuotos ir intensyviai spalvotos. Tokie skirtumai buvo vadinami heteropiknoze. Terminas "heterochromatinas" buvo pasiūlytas chromosomų sritims, rodančioms teigiamą heteropiknozę visuose mitozinio ciklo etapuose, žymėti. Yra euchromatinas - pagrindinė mitozinių chromosomų dalis, kuri vyksta įprastu sutankinimo, dekompaktavimo ciklu mitozės metu, ir heterochromatinas - chromosomų sekcijos, kurios nuolat yra kompaktiškos būsenos.
Daugumos eukariotų rūšių chromosomose yra ir eu, ir heterochromatino regionai, pastarieji sudaro didelę genomo dalį. Heterochromatinas yra centromerinėse, kartais telomerinėse srityse. Heterochromatiniai regionai buvo rasti euchromatinėse chromosomų rankose. Jie atrodo kaip heterochromatino interkalacijos (interkalacijos) į euchromatiną. Toks heterochromatinas vadinamas interkalariniu. Chromatino sutankinimas. Euchromatinas ir heterochromatinas skiriasi tankinimo ciklais. Euhr. praeina visą tankinimo-dekompaktavimo ciklą nuo tarpfazės iki tarpfazės, hetero. išlaiko santykinio kompaktiškumo būseną. Diferencinis dažymas. Skirtingos heterochromatino sekcijos nudažomos skirtingais dažais, vienos vietos – vienais, kitos – keliais. Naudojant įvairias dėmes ir naudojant chromosomų pertvarkymus, kurie pažeidžia heterochromatino sritis, buvo apibūdinta daug mažų Drosophila regionų, kuriuose afinitetas spalvai skiriasi nuo gretimų regionų.
Kariotipo samprata (apibrėžimas) Bendrosios žmogaus kariotipo charakteristikos.
Kariotipas - diploidinis chromosomų rinkinys, būdingas tam tikros rūšies organizmų somatinėms ląstelėms, kuris yra rūšiai būdingas požymis ir pasižymi tam tikru chromosomų skaičiumi, struktūra ir genetine sudėtimi.
Jei žymimas chromosomų skaičius haploidiniame lytinių ląstelių rinkinyje P, Tai bendroji formulė atrodys kariotipas 2p, kur vertė P skirtingose rūšyse skiriasi. Kadangi kariotipas yra organizmams būdinga rūšis, atskiriems individams kariotipas gali skirtis dėl tam tikrų savybių. Pavyzdžiui, skirtingų lyčių atstovuose iš esmės yra tos pačios chromosomų poros ( autosomos), bet jų kariotipai skiriasi viena chromosomų pora ( heterochromosomos, arba lytinės chromosomos). Kartais šiuos skirtumus sudaro skirtingas moterų ir vyrų heterochromosomų skaičius (XX arba XO). Dažniau skirtumai yra susiję su lytinių chromosomų sandara, žymima skirtingomis raidėmis -X ir Y (XX arba XY).
Kiekvienam kariotipo chromosomų tipui, turinčiam tam tikrą genų kompleksą, yra du homologai, paveldėti iš tėvų su jų lytinėmis ląstelėmis. Dvigubas genų rinkinys, esantis kariotipe – genotipas – yra unikalus suporuotų genomo alelių derinys. Genotipas apima konkretaus individo vystymosi programą.
Denverio (1960) ir Paryžiaus (1971) žmogaus chromosomų klasifikacija: pagrindiniai principai ir esmė.
Denverio ir Paryžiaus chromosomų klasifikacija Chromosomos skirstomos į autosomas (somatines ląsteles) ir heterochromosomas (lytines ląsteles). Levitsky (1924) siūlymu, diploidinis ląstelės somatinių chromosomų rinkinys buvo vadinamas kariotipas. Jai būdingas chromosomų skaičius, forma, dydis. Apibūdinti kariotipo chromosomas, remiantis S.G. Navashina jie yra išdėstyti forma idiogramos - sisteminis kariotipas. 1960 metais buvo pasiūlyta Denverio tarptautinė chromosomų klasifikacija, kurioje chromosomos klasifikuojamos pagal centromero dydį ir vietą. Žmogaus somatinės ląstelės kariotipe išskiriamos 22 poros autosomų ir pora lytinių chromosomų. Chromosomų rinkinys somatinėse ląstelėse vadinamas diploidas , ir lytinėse ląstelėse - haploidas (Jis lygus pusei autosomų rinkinio). Žmogaus kariotipo idiogramoje chromosomos skirstomos į 7 grupes, priklausomai nuo jų dydžio ir formos. 1 - 1-3 dideli metacentriniai. 2 - 4-5 dideli submetacentriniai. 3 - 6-12 ir X-chromosomų terpė metacentrinė. 4 - 13-15 vidutinis akrocentrinis. 5 - 16-18 santykinai mažas meta-submetacentrinis. 6 - 19-20 mažas metacentrinis. 7 - 21-22 ir Y-chromosoma yra mažiausia akrocentrinė. Pagal Paryžiaus klasifikacija chromosomos skirstomos į grupes pagal dydį ir formą, taip pat tiesinę diferenciaciją.