Учебное пособие предназначено для студентов направления «Биология» всех профилей подготовки, всех форм обучения для теоретической подготовки к занятиям, зачетам и экзаменам. Пособие охватывает основные разделы структурной биохимии: строение, физико-химические свойства и функции основных классов биологических макромолекул. Большое внимание уделено ряду прикладных аспектов биохимии.
Нуклеотиды и нуклеиновые кислоты
Структура нуклеотидов и азотистых оснований
Нуклеотиды принимают участие во множестве биохимических процессов, а также являются мономерами нуклеиновых кислот. Нуклеиновые кислоты обеспечивают все генетические процессы. Каждый нуклеотид состоит из трех типов химических молекул:
Азотистое основание;
Моносахарид;
1-3 остатка фосфорной кислоты.
В отличие от моносахаридов, нуклеотиды как мономеры являются сложно устроенными молекулами, состоящими из структур, относящихся к разным классам химических веществ, поэтому необходимо рассмотреть свойства и структуру этих компонентов по отдельности.
Азотистые основания
Азотистые основания относятся к гетероциклическим соединениям. В состав гетероцикла помимо атомов углерода входят атомы азота. Все азотистые основания, входящие в нуклеотиды относят к двум классам азотистых оснований: пуриновые и пиримидиновые. Пуриновые основания это производные пурина – гетероцикла, состоящего из двух циклов, один пятичленный, второй – шести, нумерация осуществляется так, как показано на рисунке. Пиримидиновые основания являются производными пиримидина и состоят из одного шестичленного цикла, нумерация также указана на рисунке (Рисунок 31). Главные пиримидиновые основания и у прокариот, и у эукариот – это цитозин, тимин и урацил. Из пуриновых оснований чаще всего встречаются аденин и гуанин. Два других – ксантин и гипоксантин – являются интермедиатами в процессах их метаболизма. У человека в роли конечного продукта катаболизма пуринов выступает окисленное пуриновое основание – мочевая кислота . Помимо пяти названных выше главных оснований известны и менее широко представленные минорные основания. Некоторые из них присутствуют только в нуклеиновых кислотах бактерий и вирусов, но многие также найдены в составе про- и эукариотических ДНК и транспортных и рибосомных РНК. Так, и бактериальная ДНК, и ДНК человека содержат значительные количества 5-метилцитозина; в бактериофагах обнаружен 5-гидроксиметилцитозин. Необычные основания выявлены в матричной РНК – N 6 -метиладенин, N 6 , N 6 -диметиладенин и N 7 -Meтилгуанин. У бактерий также обнаружен модифицированный урацил с присоединенной по N 3 -положению (α-амино, α-карбокси) -пропильной группой. Функции этих замещенных пуринов и пиримидинов до конца не выяснены, однако они могут образовывать неканонические связи между основаниями (это будет рассмотрено ниже), обеспечивая образование вторичных и третичных структур нуклеиновых кислот.
Рисунок 31. Структура азотистых оснований
В клетках растений выявлена серия пуриновых оснований с метильными заместителями. Многие из них фармакологически активны. В качестве примера можно привести кофейные зерна, содержащие кофеин (1,3, 7-триметилксантин), чайный лист, содержащий теофиллин (1, 3-диметил-ксантин), и какао-бобы, в состав которых входит теобромин (3, 7-диметилксантин).
изомерия и Физико-химические свойства пуриновых и пиримидиновых оснований
Молекула азотистого основания образует систему чередующихся одинарных и двойных связей (систему сопряженных двойных связей). Такая организация образует жесткую молекулу, без возможности конформационных переходов. В результате нельзя говорить об изменении конформации азотистых оснований.
Для азотистых оснований выявлен только один тип изомерии кето-енольный переход или таутомерия.
Таутомерия
Благодаря феномену кето-енольной таутомерии нуклеотиды могут существовать либо в лактимной, либо в лактамной формах, причем в физиологических условиях лактамная форма превалирует у гуанина и тимина (Рисунок 32). Важность этого обстоятельства станет ясна при обсуждении процессов спаривания оснований.
Рисунок 32. Таутомерия нуклеотидов
Растворимость
При нейтральном рН наименьшей растворимостью обладает гуанин. Следующим в этом ряду стоит ксантин. Мочевая кислота в форме уратов сравнительно неплохо растворяется при нейтральном рН, но очень плохо растворима в жидкостях с более низкими значениями рН, таких, как моча. Гуанин в моче человека в норме отсутствует, а ксантин и мочевая кислота являются ее обычными компонентами. Последние два пурина часто входят в состав камней мочевого тракта.
Поглощение света
За счет системы сопряженных двойных связей все азотистые основания поглощают в ультрафиолетовой части спектра. Спектр поглощения – график распределения оптической плотности в зависимости от длины волны. Для каждого азотистого основания свой спектр поглощения, по нему можно различить растворы различных азотистых оснований или соединений в состав которых входит азотистое основание (нуклеотиды), но максимум поглощения у всех совпадает при длине волны 260 нм. Это позволяет легко и быстро определять концентрацию как азотистых оснований, так нуклеотидов и нуклеиновых кислот. Спектр поглощения также зависит от рН раствора (Рисунок 33).
Рисунок 33. Спектры поглощения различных азотистых оснований
Функции азотистых оснований
Азотистые основания практически не встречаются в свободном состоянии. Исключение составляют некоторые алкалоиды и мочевая кислота.
Азотистые основания выполняют следующие функции:
Входят в состав нуклеотидов;
Часть алкалоидов – азотистые основания, например, кофеин в кофе или теофелин в чае;
Промежуточные продукты обмена азотистых оснований и нуклеотидов;
Мочевая кислота – причина мочекаменной болезни;
В виде мочевой кислоты выводится азот у некоторых организмов.
Нуклеотиды и нуклеозиды
Молекулы нуклеозидов построены из пуринового или пиримидинового основания, к которому (β-связью присоединен углевод (обычно D-рибоза или 2-дезоксирибоза) в N 9 или N 1 ‒положении соответственно. Таким образом, адениновый рибонуклеозид (аденозин) состоит из аденина и D-рибозы, присоединенной в положении N 9 ; гуанозин – из гуанина и D-рибозы в положении N 9 ; цитидин – из цитозина и рибозы в положении N 1 ; уридин – из урацила и рибозы в положении N 1 . Таким образом в пуриновых нуклеозидах (нуклеотидах) азотистое основание и сахар связаны 1-9 β гликозидной связью, а в пиримидинах – 1-1 β гликозидной связью.
В состав 2́-дезоксирибонуклеозидов входят пуриновые или пиримидиновые основания и 2́-дезоксирибоза, присоединенная по тем же атомам N 1 и N 9 . Присоединение рибозы или 2́-дезоксирибозы к кольцевой структуре основания происходит за счет относительно кислотолабильной N-гликозидной связи (Рисунок 34).
Нуклеотиды – это производные нуклеозидов, фосфорилированные по одной или более гидроксильным группам остатка рибозы (или дезоксирибозы). Так, аденозинмонофосфат (AMФ или аденилат) построен из аденина, рибозы и фосфата. 2́-дезоксиаденозинмонофосфат (дAMФ или дезоксиаденилат) представляет собой молекулу, состоящую из аденина, 2́-дезоксирибозы и фосфата. Обычно к урацилу присоединена рибоза, к тимину – 2́-дезоксирибоза. Поэтому тимидиловая кислота (ТМФ) состоит из тимина, 2́-дезоксирибозы и фосфата. Кроме вышеперечисленных форм нуклеотидов обнаружены и нуклеотиды необычной структуры. Так, в молекуле тРНК выявлен нуклеотид, в котором рибоза присоединяется к урацилу в пятом положении, т. е. не азот-углеродной связью, а углерод-углеродной. Продукт этого необычного присоединения назван псевдоуридином (ψ). Молекулы тРНК содержат и другую необычную нуклеотидную структуру – тимин, соединенный с рибозомонофосфатом. Этот нуклеотид образуется уже после синтеза молекулы тРНК путем метилирования остатка УMФ S-аденозилметионином. Псевдоуридиловая кислота (ψМФ) тоже образуется в результате перегруппировки УMФ после синтеза тРНК.
Рисунок 34. Структура пуриновых и пиримидиновых нуклеозидов и нуклеотидов
Номенклатура, физико-химические свойства и функции нуклеозидов и нуклеотидов
Положение фосфатной группы в молекуле нуклеотида указывается цифрой. Например, аденозин с фосфатной группой, присоединенной к 3-му углероду рибозы, должен быть обозначен как 3́-монофосфат. Штрих после цифры ставят для того, чтобы отличить номер углерода в пуриновом или пиримидиновом основании от положения этого атома в остатке дезоксирибозы. При нумерации атомов углерода основания штрих не ставится. Нуклеотид 2́-дезоксиаденозин с фосфатным остатком при углероде-5 молекулы сахара обозначается как 2́-дезоксиаденозин-5́-монофосфат. Нуклеозиды, содержащие аденин, гуанин, цитозин, тимин и урацил, принято обозначать буквами A, Г, Ц, Т и У соответственно. Наличие буквы d (или д) перед сокращением обозначает, что углеводным компонентом нуклеозида является 2́-дезоксирибоза. Гуанозин, содержащий 2́-дезоксирибозу, может быть обозначен дГ (дезоксигуанозин), а соответствующий ему монофосфат с фосфатной группой, присоединенной к третьему атому углерода дезоксирибозы, – дГ-3́-МФ. Как правило, в тех случаях, когда фосфат присоединен к углероду-5 рибозы или дезоксирибозы, символ 5́ опускается. Так, гуанозин 5́-монофосфат принято обозначать ГМФ, а 5́-монофосфат 2́-дезоксигуанозина сокращают как дГМФ. Если к углеводному остатку нуклеозида присоединены 2 или 3 остатка фосфорной кислоты используются аббревиатуры ДФ (дифосфат) и ТФ (трифосфат). Таким образом, аденозин + трифосфат с тремя фосфатными группами в 5́-положении углевода будет обозначаться АТФ. Поскольку в молекулах нуклеотидов фосфаты находятся в виде ангидридов фосфорной кислоты, т. е. в состоянии с низкой энтропией, их называют макроэргами (обладающими большим запасом потенциальной энергии). При гидролизе 1 моля АТФ до AДФ высвобождается 7,3 кКал потенциальной энергии.
Рисунок 35. Структура цАМФ
Физико-химические свойства нуклеотидов
Так как в состав нуклеотидов входят азотистые основания, то такие свойства как таутомерия и способность поглощать в ультрафиолетовой части спектра также характерны и для нуклеотидов, причем спектры поглощения азотистых оснований и содержащих эти основания нуклеотидов сходны. Наличие сахара и остатков фосфорной кислоты делает их более гидрофильными чем азотистые основания. Все нуклеотиды являются кислотами, так как содержат остатки фосфорной кислоты.
Функции природных нуклеотидов
Нуклеотиды являются мономерами нуклеиновых кислот (РНК, ДНК). В состав ДНК входят дезоксирибонуклеотидфосфаты – производные аденина, тимина, гуанина и цитозина. Также некоторые молекулы гуанина и цитозина в составе ДНК метилированы, то есть содержат метильную группу. Как основные мономеры в состав РНК входят рибонуклеотидфосфаты – производные аденина, урацила, гуанина и цитозина. Также в состав РНК входят нуклеотиды, содержащие различные минорные азотистые основания, например ксантин, гипоксантин, дигидроуридин и др.
Нуклеотиды являются мономерами коферментов (НАД, НАДФ, ФАД, ко-энзим А, метионин-аденозин). В составе коферементов они участвуют в ферментативных реакциях. Более подробно эта функция будет рассмотрена ниже.
Энергетическая (АТФ) . АТФ выполняет функцию основного внутриклеточного переносчика свободной энергии. Концентрация наиболее распространенного свободного нуклеотида в клетках млекопитающих – АТФ – составляет около 1 ммоль/л.
Сигнальная (цГМФ, цАМФ) (Рисунок 35). Циклический AMФ (3́-, 5́-аденозинмонофосфат, цАМФ) – медиатор различных внеклеточных сигналов в клетках животных – образуется из АТФ в результате реакции, катализируемой аденилатциклазой. Активность аденилатциклазы регулируется комплексом взаимодействий, многие из которых инициируются через рецепторы гормонов. Внутриклеточная концентрация цАМФ (около 1 мкмоль/л) на 3 порядка ниже концентрации ATФ. Циклический цГМФ (3́-, 5́-гуанозинмонофосфат, цГМФ) служит внутриклеточным проводником внеклеточных сигналов. В некоторых случаях цГМФ выступает в роли антагониста цАМФ. цГМФ образуется из ГТФ под действием гуанилатциклазы – фермента, имеющего много общего с аденилатциклазой. Гуанилатциклаза, как и аденилатциклаза, регулируется различными эффекторами, в том числе и гормонами. Как и цАМФ, цГМФ гидролизуется фосфодиэстеразой до соответствующего 5́-монофосфата.
Регуляторная (ГТФ) . Активность группы белков (G-белков), выполняющих в основном регуляторную функцию, зависит от того: какой нуклеотид они связывают. В неактивной форме эти белки связывают ГДФ, при активации белка происходит замена ГДФ на ГТФ. При выполнении своей функции белок гидролизует ГТФ до ГДФ и фосфата, выделившаяся, энергия затрачивается на функционирование белка.
Активация при метаболизме липидов и моносахаридов (УТФ, СТФ) . Производные урациловых нуклеотидов участвуют в качестве активирующих агентов в реакциях метаболизма гексоз и полимеризации углеводов, в частности при биосинтезе крахмала и олигосахаридных фрагментов гликопротеинов и протеогликанов. Субстратами в этих реакциях являются уридин-дифосфатсахара. Например, уридиндифосфатглюкоза служит предшественником гликогена. Также превращение глюкозы в галактозу, глюкуроновую кислоту или другие производные моносахаридов происходит в виде коньюгата с УДФ. СТР необходим для биосинтеза некоторых фосфоглицеридов в тканях животных. Реакции с участием церамида и ЦДФ-холина приводят к образованию сфингомиелина и других замещенных сфингозинов.
Участие в дезактивации различных спиртов и фенолов (УДФ-глюкуроновая кислота). Уридиндифосфатглюкуроновая кислота – выполняет функцию «активного» глюкуронида в реакциях конъюгирования, например, при образовании глюкуронида билирубина.
Нуклеотиды в составе коферментов
Коферменты – это низкомолекулярные соединения связанные с ферментами (см раздел «Ферменты») непосредственно участвующие в в биохимической реакции, другими словами это еще один субстрат, не выходящий в окружающую среду.
Коферменты подразделяют на две группы:
переносчики протонов и электронов, эти коферменты участвуют в окислительно-восстановительных реакциях;
переносчики всех остальных групп кроме протонов и электронов, эти коферменты участвуют в трансферазных реакциях.
Более подробно механизмы упомянутых реакций можно рассмотреть в главе «Ферменты».
Некоторые коферменты содержат в своем составе нуклеотиды. Они также делятся на эти же две группы.
Коферменты переносчики протонов и электронов
Эти коферменты участвуют в окислительно-восстановительных реакциях, где аденозин выполняет только структурную функцию, в реакцию вступают нуклеотиды, содержащие другие типы оснований, выделяют два типа таких коферментов: никотиновые и флавиновые. Они отличаются не только по активной группировке, но и по типу реакций, которые они осуществляют.
Никотиновые коферменты
Рисунок 36. Никотиновые коферменты. А-структура NAD, Б-структура NADP, В-механизм активности никотиновой кислоты, Г-механизм работы никотиновых коферментов
Никотинамидадениндинуклеотид (NAD +) – главный акцептор электронов при окислении топливных молекул. Реакционноспособная часть NAD + – его никотинамидное кольцо. При окислении субстрата никотинамидное кольцо NAD + присоединяет ион водорода и два электрона, которые являются эквивалентами гидрид-иона. Восстановленная форма этого переносчика – NADH. В ходе этого дегидрирования один атом водорода субстрата прямо переносится на NAD + , тогда как второй переходит в растворитель. Оба электрона, теряемые субстратом, переносятся на никотинамидное кольцо. Роль донора электронов в большинстве процессов восстановительного биосинтеза (пластического обмена); выполняет восстановленная форма никотин амидадениндинуклеотидфосфата (NADPH). NADPH отличается от NAD наличием фосфата, связанного эфирной связью с 2́-гидроксильной группой аденозина. Окисленная форма NADPH обозначается как NADP + . NADPH переносит электроны таким же образом, как NADH. Однако, NADPH используется почти исключительно в процессах восстановительного биосинтеза, тогда как NADH используется преимущественно для генерирования АТР. Дополнительная фосфатная группа NADPH – это участок, ответственный за осуществление целевого предназначения молекулы, состоящего в распознавании ферментами.
Флавиновые коферменты
Первый флавиновый кофермент (флавинмононуклеотид FMN) был выделен А. Сент-Дьёрдьи из сердечной мышцы в 1932 г., Р. Г. Варбург и В. Христиан тогда же получили из дрожжей первый флавопротеид, содержащий FMN в качестве кофермента. Второй важнейший флавиновый кофермент – флавинадениндинуклеотид (FAD) выделен ими же как кофактор оксидазы D-аминокислот в 1938 году. За счет окислительно-восстановительного превращения флавинового кольца флавиновые коферменты осуществляют окислительно-восстановительные реакции в составе многих важнейших ферментных систем: оксидаз (в частности, оксидаз D- и L-аминокислот, моноаминооксидазы, регулирующей уровень катехоламинов в крови) и дегидрогеназ (часто с участием никотинамидадениндинуклеотида и убихинонов).
Рисунок 37. Флавиновые коферменты. А-структура FAD, Б-механизм активности никотиновой кислоты, В-механизм работы флавиновых коферментов
Второй основной переносчик электронов при окислении топливных молекул – флавинадениндинуклеотид. Сокращения, используемые для обозначения окисленной и восстановленной форм этого переносчика – соответственно FAD и FADH 2 . Реакционноспособная часть FAD – это его изоаллоксазиновое кольцо. FAD, подобно NAD + , присоединяет два электрона. Однако FAD в отличие от NAD + присоединяет оба теряемых субстратом атома водорода.
Конец ознакомительного фрагмента.
Молекула нуклеотида имеет в своем составе сахар, фосфат и азотистую основу. Как эти простые компоненты позволяют нуклеотидам объединяться вместе, чтобы создавать такие полимеры, как ДНК и РНК, а также молекулы, несущие энергию, такие как АТФ?
Нуклеотиды: часть структуры ДНК
Что такое нуклеотид? Чтобы это понять, нужно представить себе ДНК. Попав в ядро клетки и распутав хромосомы, можно увидеть тонкую двойную нить. При масштабировании можно увидеть, что каждая из этих нитей состоит из небольших строительных блоков, называемых нуклеотидами.
Если ДНК выглядит как скрученная лестница, каждый строительный блок или нуклеотид включает половину ступени и немного вертикальной части лестницы. Другая половина ступени относится к соседней цепочке ДНК. Нуклеотиды также могут существовать сами по себе или быть частью других важных молекул, помимо ДНК. Например, энергетический носитель АТФ представляет собой форму нуклеотида.

Компоненты нуклеотида
В состав нуклеотида входят такие компоненты, как азотистая основа, сахар и один или несколько фосфатов. Стоит рассмотреть каждый их них более подробно:
- Азотистое основание. Это может быть аденин, тимин, цитозин, гуанин, урацил. Они не являются кислотами, каждый из них содержит несколько атомов азота. Нуклеотиды могут соединяться друг с другом: цитозин всегда составляет пару с гуанином и адениновые пары с тимином в ДНК или урацил в РНК.
- Следующим основным компонентом нуклеотида является сахар. Существует много видов сахара, но здесь важны два: рибоза - это сахар, который вы увидите в РНК. Существует версия рибозы, у которой отсутствует атом кислорода, и он будет называться сахарной дезоксирибозой. Это тип сахара в ДНК-нуклеотидах. Помните, что ДНК - это дезоксирибонуклеиновая кислота.
- Последним основным фрагментом нуклеотида является фосфат. Фосфат представляет собой атом фосфора, связанный с четырьмя атомами кислорода. Связи между фосфатами являются очень высокой энергией и действуют как форма хранения энергии. Когда связь сломана, полученная энергия может быть использована для выполнения работы.

Типы нуклеотидов
Когда нуклеотиды полимеризуются или объединяются вместе, они образуют нуклеиновую кислоту, такую как ДНК или РНК. Каждый нуклеотидный фосфат присоединяется к другому сахару, образуя сахар-фосфатную основу с азотистыми основаниями. Нуклеозид является частью нуклеотида, который состоит только из сахара и основания. Таким образом, мы можем говорить о нуклеотиде как о нуклеозиде и фосфатах:
- Нуклеозид монофосфат представляет собой нуклеотид, который включает в себя один фосфат.
- Нуклеозид дифосфат представляет собой нуклеотид, который включает в себя два фосфата.
- Нуклеозид трифосфат представляет собой нуклеотид, который содержит три фосфата. Нуклеотиды являются строительными блоками ДНК и РНК.
Какие различают типы нуклеотидов, какова их структура и как изменение одного нуклеотида может повлиять на выживание организма?

Нуклеотид - это в биологии... (определение)
ДНК человека состоит из нуклеотидов, которые в основном представляют собой субэлементное измерение ДНК, выстраиваемое парами. Есть около 3 миллиардов этих пар, также называемых парами оснований. Какое можно дать определение нуклеотиду? Каждый сперматозоид и каждая яйцеклетка содержат примерно шесть миллиардов отдельных нуклеотидов в своем ядре, которые организованы в компактные молекулы ДНК. Это облегчает их хранение и перемещение.
Итак, что такое нуклеотиды? Они действуют как особый язык, который используется для написания рецептов химических веществ, создаваемых вашим организмом, в частности белков. Большинство участков нуклеотидов называют нежелательной ДНК, потому что они ничего не кодируют. Тем не менее есть небольшая доля, которая имеет решающее значение для вашего выживания и делает вас такими, какие вы есть. Этот 2 % кода нуклеотидов для каждого белка, который ваш организм производит и имеет на участках ДНК, называемых генами. Каждый ген кодирует цепь аминокислот, которая приводит к образованию определенного белка.
Мутации, которые являются изменениями в ДНК-клетки, с участием одного нуклеотида, могут показаться тривиальными, учитывая, что в геноме человека так много нуклеотидов, но, когда они происходят на определенных генах, они могут привести к опасным для жизни заболеваниям. Чтобы лучше понять этот механизм, нужно сначала взглянуть на некоторые основы нуклеотидов.
Структура нуклеотидов
Нуклеотиды представляют собой мономеры (или строительные блоки) нуклеиновых кислот и состоят из 5-углеродного сахара, фосфатной группы и азотистого основания. Как уже было сказано, сахар и основание вместе образуют нуклеозид. Добавление фосфатной группы превращает молекулу в нуклеотид. Нуклеотиды называются в соответствии с азотистым основанием, которое они содержат, и сахаром, присоединенным к нему (например, дезоксирибозой в ДНК-нуклеотидах и рибозе в РНК). Какие нуклеотиды в ДНК и РНК? Всего существует восемь различных нуклеозидов в ДНК и РНК:
- РНК: аденозин, гуанозин, цитидин, уридин.
- ДНК: дезоксиаденозин, дезоксигуанозин, деоксицитидин, дезокситимидин.
Существуют и другие важные нуклеотиды, такие как те, которые участвуют в метаболизме (например, АТФ) и клеточной передаче сигналов (например, ГТФ).

Связывание нуклеотидов
Для создания цепей полимера (или нескольких единиц), которые приводят к образованию РНК и ДНК, нуклеотиды соединяются друг с другом через сахарофосфатный скелет, который образуется, когда фосфат одного нуклеотида присоединяется к сахару другого. Это возможно благодаря сильным ковалентным связям, называемым фосфодиэфирными связями.
Поскольку ДНК представляет собой двухцепочечную молекулу, две из этих полимерных цепей должны присоединяться друг к другу, как лестница. «Ступеньки» состоят из пар нуклеотидов, которые соединяют две стороны лестницы с помощью водородных связей. Что такое нуклеотид? Это структурная единица ДНК, которая состоит из азотистого основания и сахар-фосфатной основной цепи, состоящей из фосфатной группы и сахара. ДНК состоит из многих нуклеотидов, которые содержат и защищают генетические коды организма.

Нуклеиновые кислоты
Нуклеиновые кислоты являются биополимерами, которые наряду с белками играют важную роль в клетках всех живых организмов. Эти соединения ответственны за хранение, передачу и реализацию наследственной информации. Что такое нуклеотиды? Это мономеры нуклеиновых кислот.
Между частями нуклеотида возникают ковалентные химические связи, которые образуются в результате реакций конденсации. Такие реакции являются обратными гидролизу. Интересным фактом является то, что молекулы ДНК обычно не только длиннее, чем молекулы РНК, но и включают в себя две цепочки, которые соединены друг с другом при помощи водородных связей, возникающих между азотистыми основаниями.
- это сложные мономеры, из которых собраны гетерополимерные молекулы. ДНК и РНК. Свободные нуклеотиды участвуют в сигнальных и энергетических процессах жизнедеятельности. ДНК-нуклеотиды и РНК-нуклеотиды имеют общий план строения, но различаются по строению сахара-пентозы. В ДНК-нуклеотидах используется сахар дезоксирибоза, а в РНК-нуклеотидах - рибоза.
Структура нуклеотида
В каждом нуклеотиде можно выделить 3 части:
1. Углевод - это пятичленный сахар-пентоза (рибоза или дезоксирибоза).
2. Фосфорный остаток (фосфат) - это остаток фосфорной кислоты.
3. Азотистое основание - это соединение, в котором много атомов азота. В нуклеиновых кислотах используется всего 5 видов азотистых оснований: Аденин, Тимин, Гуанин, Цитозин, Урацил. В ДНК - 4 вида: Аденин, Тимин, Гуанин, Цитозин. В РНК - тоже 4 вида: Аденин, Урацил, Гуанин, Цитозин, Легко заметить, что в РНК происходит замещение Тимина на Урацил по сравнению с ДНК.
Общая структурная формула пентозы (рибозы или дезоксирибозы), молекулы которой образуют "скелет" нуклеиновых кислот:
Если Х заменить на Н (Х = Н) - то получаются дезоксирибонуклеозиды; если Х заменить на ОН (Х = ОН) - то получаются рибонуклеозиды. Если вместо R подставить азотистое основание (пуриновое или пиримидиновое) - то получится конкретный нуклеотид.
Важно обратить внимание на те положения атомов углерода в пентозе, которые обозначены как 3" и 5". Нумерация атомов углерода начинается от атома кислорода вверху и идёт по часовой стрелке. Последним получается атом углерода (5"), который располагается за пределами пентозного кольца и образует, можно сказать, "хвостик" у пентозы. Так вот, при наращивании цепочки из нуклеотидов фермент может присоединить новый нуклеотид только к углероду 3" и ни к какому другому. Поэтому 5"-конец нуклеотидной цепочки никогда не сможет иметь продолжения, удлинняться может только 3"-конец.



Сравните нуклеотид для РНК с нуклеотидом для ДНК.
Попробуйте узнать, какой это нуклеотид, в таком представлении:

АТФ - свободный нуклеотид
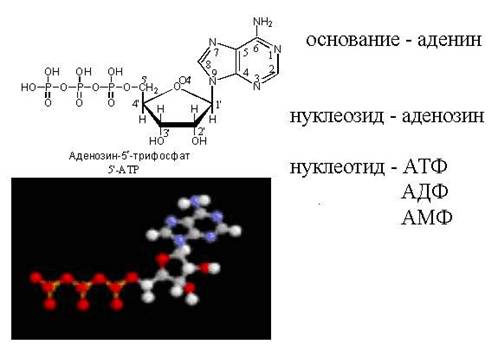
цАМФ - "закольцованная" молекула АТФ

Схема строения нуклеотида

Обратите внимание на то, что активированный нуклеотид, способный наращивать цепочку ДНК или РНК, имеет "трифосфатный хвостик". Именно этим "энергонасыщенным" хвостиком он может присоединиться к уже имеющейся цепочке растущей нуклеиновой кислоты. Фосфатный хвостик сидит на 5-м атоме углерода, так что это положение углерода уже занято фосфатами и предназнено для прикрепления. К чему же его прикрепить? Только к углероду в положении 3". После прикрепления данный нуклеотид сам станет мишенью дла прикрепления следующего нуклеотида. "Принимающая сторона" предоставляет углерод в положении 3", а "прибывающая сторона" цепляется к нему фосфатным хвостиком, находящимся в положении 5". В целом цепочка растёт со стороны 3".
Наращивание нуклеотидной цепочки ДНК
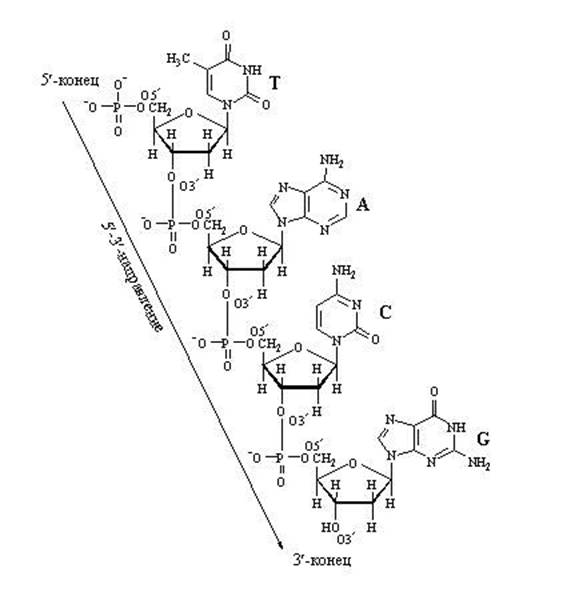
Наращивание цепочки за счёт "продольных" связей между нуклеотидами может идти только в одном направлении: от 5" ⇒ к 3", т.к. новый нуклеотид можно присоединить только к 3"-концу цепочки, но не к 5"-концу.
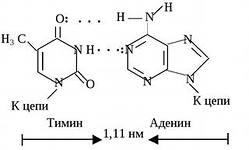
Пары нуклеотидов, связанные "поперечными" комплементарными связями своих азотистых оснований

Участок двойной спирали ДНК
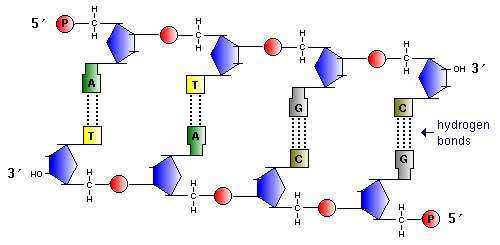
Найдите признаки антипараллельности двух цепей ДНК.
Найдите пары нуклеотидов с двойными и тройными комплементарными связями.
Нуклеотиды
Остановимся подробнее на нуклеотидах. Известно, что нуклеотиды называются аденин, гуанин, тимин, цитозин и урацил – азотистые основания, они представлены на рисунке ниже.
Нуклеотиды – это мономеры нуклеиновых кислот. Нуклеиновые кислоты в эукариотических клетках находятся в ядре. Они есть у всех живых организмов (у тех, у кого нет ядра, нуклеиновые кислоты все равно есть – они находятся в центре клетки у бактерий и образуют нуклеоиды). Мономеры, из которых потом строятся нуклеиновые кислоты, состоят из азотистого основания, остатка сахара (дезоксирибоза или рибоза) и фосфата. Сахара вместе с азотистым основанием называются нуклеозидами (аденозин, гуанозин, тимидин, цитидин). Если к ним присоединены 1-, 2-, или 3-фосфорных остатка, то вся эта структура называется Соответственно, нуклеотизид монофосфатом, дифосфатом или трифосфатом или нуклеотидом (аденин, гуанин, тимин, цитозин).
Вот так модель АТФ выглядит в пространстве. Азотистое основание, входящее в состав ДНК делится на две группы – пиримидиновую и пуриновую. В состав ДНК входит аденин, тимин, цитозин и гуанин, в РНК вместо тимина урацил. Как известно, ДНК – это большой архив, в котором хранится информация, а РНК – это молекула, которая переносит информацию из ядра в цитоплазму для синтеза белков. С различием в функциях связаны различия в строении. РНК более химически активно из-за того, что ее сахар - рибоза – имеет в своем составе гидроксильную группу, а в дезоксирибозе кислорода нет. Из-за отсутствия кислорода ДНК более инертно, что важно для ее функции хранения информации, чтобы она не вступала ни в какие реакции.
Нуклеотиды способны взаимодействовать друг с другом, при этом «выбрасывается» два фосфора, и между соседними нуклеотидами образуется связь. В молекуле фуранозы молекулы углерода пронумерованы. С первым связано азотистое основание. Когда образуется цепочка нуклеотидов, связь осуществляется между пятым углеродом одной и третьим углеродом другой фосфорной кислоты. Поэтому в цепочке нуклеиновых кислот выделяют разные неравнозначные концы, относительно которых молекула не симметрична.
Комплементарные друг другу одноцепочечные молекулы нуклеиновой кислоты способны образовывать двуцепочечную структуру. Внутри этой спирали аденин образует пару с тимином, а гуанин - с цитозином. Встречается утверждение, что нуклеотиды подходят друг другу как осколки разбитого стекла, поэтому они и образуют пары. Но это утверждение неверно. Нуклеотиды способны образовывать пары как угодно. Единственная причина, по которой они соединяются так, и никак иначе, заключается в том, что угол между «хвостиками», которые идут к сахарам, совпадает только в этих парах, и, кроме того, совпадают их размеры. Никакая другая пара не образует такой конфигурации. А поскольку они совпадают, то их через сахаро-фосфатный остов можно связать друг с другом. Структуру двойной спирали открыли в 1953 году Джеймс Уотсон и Фрэнсис Крик.
При соединение друг с другом против 5’-конца одной нити находится 3’-конец другой нити. То есть нити идут в противоположных направлениях – говорят, что нити в ДНК антипараллельны.
На рисунке видна модель ДНК, видно, что аденин соединяется с тимином двумя водородными связями, а гуанин соединяется с цитозином тройной водородной связью. Если молекулу ДНК подогревать, то ясно, что две связи легче разорвать, чем три, это существенно для свойств ДНК.
В силу пространственного расположения сахаро-фосфатного остова и нуклеотидов, когда нуклеотиды накладывают один на другой и «сшивают» через сахаро-фосфатный остов, цепочка начинает заворачиваться, тем самым образуя знаменитую двойную спираль.
На рисунках представлены шариковые модели ДНК, где каждый атом обозначен шариком. Внутри спирали имеются бороздки: маленькая и большая. Через эти бороздки с ДНК взаимодействуют белки и распознают там последовательность нуклеотидов.
При нагревании ДНК водородные связи разрываются и нити в двойной спирали расплетаются. Процесс нагревания называется плавлением ДНК, при этом разрушаются связи между парами А-Т и Г-Ц.Чем больше в ДНК пар А-Т, тем менее прочно нити друг с другом связаны, тем легче ДНК расплавить. Переход из двухспиральной ДНК в одно-спиральную измеряется на спектрофотометрах по поглощению света при 260 нм. Температура плавления ДНК зависит от А-Т/Г-Ц состава и размера фрагмента молекулы. Ясно, что если фрагмент состоит из нескольких десятков нуклеотидов, то его гораздо легче расплавить, чем более длинные фрагменты.
У человека в гаплоидном геноме, то есть единичном наборе хромосом, 3 млрд. пар нуклеотидов, и их длина составляет 1,7 м, а клетка гораздо меньше, как вы догадываетесь. Для того, чтобы ДНК смогла в ней поместиться, она достаточно плотно свернута, и в эукариотической клетке свернуться ей помогают белки – гистоны. Гистоны имеют положительный заряд, а так как ДНК заряжена отрицательно, то гистоны обладают сродством к ДНК. Упакованная при помощи гистонов ДНК имеет вид бусин, называемых нуклеосомами. 200 пар нуклеотидов идет на одну нуклеосому, 146 пар накручиваются на гистоны, а остальные 54 висят в виде линкерных (связывающих нуклеосомы) ДНК. Это первый уровень компактизации ДНК. В хромосомах ДНК свернута еще несколько раз для того, чтобы образовались компактные структуры.
К нуклеиновым кислотам кроме ДНК относится также РНК. В клетке присутствуют разные типы РНК: рибосомные, матричные, транспортные. Существуют и другие виды РНК, о которых мы будем говорить позже. РНК синтезируется в виде одно-цепочечной молекулы, но отдельные ее участки входят в состав двуцепочечных спиралей. Для РНК также говорят о первичной структуре (последовательности нуклеотидов) и вторичной структуре (образование двуспиральных участков).
Липиды
В состав липидов входят жирные кислоты, имеющие длинные углероводородные цепи. Жирные кислоты гидрофобны, то есть не растворимы в воде.
Липиды представляют собой соединения жирных кислот с глицерином (эфиры). Например, на рисунке изображен лецитин.
В клетке важную роль играют липиды, в которых к глицерину присоединен остаток фосфорной кислоты и 2 жирных кислоты. Они называются фосфолипидами. Молекулы фосфолипидов имеют полярную (то есть гидрофильную, хорошо растворимую) группу на одном конце молекулы и длинный гидрофобный хвост. К фосфолипидам относится фосфатидилхолин.
В водном растворе фосфолипиды образуют мицеллы, в которых молекулы обращены полярными "головами" наружу, в сторону воды, а гидрофобные "хвосты" оказываются внутри мицеллы, спрятанными от воды. Клеточную мембрану также липиды с полярными "головами", которые обращены наружу по обе стороны мембраны, а гидрофобные "хвосты" находятся внутри липидного бислоя.
Список литературы
Для подготовки данной работы были использованы материалы с сайта http://bio.fizteh.ru
К 1944 г. О. Эйвери и его коллеги К. Маклеод и М. Маккарти открыли трансформирующую активность ДНК у пневмококков. Эти авторы продолжили работу Гриффита, описавшего феномен трансформации (передачи наследственных признаков) у бактерий. О. Эйвери, К. Маклеод, М. Маккарти показали, что при удалении белков, полисахаридов и РНК трансформация бактерий не нарушается, а при воздействии на индуцирующее вещество ферментом дезоксирибонуклеазой трансформирующая активность исчезает.
В этих экспериментах впервые была продемонстрирована генетическая роль молекулы ДНК. В 1952 г. А. Херши и М. Чейз подтвердили генетическую роль молекулы ДН К в опытах на бактериофаге Т2. Пометив его белок радиоактивной серой, а ДНК-радиоактивным фосфором,они инфицировали этим бактериальным вирусом кишечную палочку Е. coli. В потомстве фага было выявлено большое количество радиоактивного фосфора и лишь следы S. Отсюда следовало, что именно ДНК, а не белок фага проникает в бактерию, а затем после репликации передается фаговому потомству.
Строение нуклеотида ДНК. Типы нуклеотидов.
Нуклеотид ДНК состоит из
Азотистого основания (в ДНК 4 типа: аденин, тимин, цитозин, гуанин)
Моносахара дезоксирибозы
Фосфорной кислоты
Молекула нуклеотида состоит из трех частей - пятиуглеродного сахара, азотистого основания и фосфорной кислоты.
Сахар, входящий в состав нуклеотида , содержит пять углеродных атомов, т. е. представляет собой пентозу. В зависимости от вида пентозы, присутствующей в нуклеотиде, различают два типа нуклеиновых кислот - рибонуклеиновые кислоты (РНК), которые содержат рибозу, и дезоксирибонуклеиновые кислоты (ДНК), содержащие дезоксирибозу. В дезоксирибозе - ОН-группа при 2-м атоме углерода заменена на атом Н, т. е. в ней на один атом кислорода меньше, чем в рибозе.
В обоих типах нуклеиновых кислот содержатся основания четырех разных видов: два из них относятся к классу пуринов и два - к классу пиримидинов. Основной характер этим соединениям придает включенный в кольцо азот. К числу пуринов относятся аденин (А) и гуанин (Г), а к числу пиримидинов - цитозин (Ц) и тимин (Т) или урацил (У) (соответственно в ДНК или РНК). Тимин химически очень близок к урацилу (он представляет собой 5-метилурацил, т. е. урацил, в котором у 5-го углеродного атома стоит метильная группа). В молекуле пуринов имеется два кольца, а в молекуле пиримидинов - одно.
Нуклеотиды соединяются между собой прочной ковалентной связью через сахар одного нуклеотида и фосфорную кислоту другого. Получаетсяполинуклеотидная цепь . На одном ее конце – свободная фосфорная кислота (5’-конец), на другом – свободный сахар (3’-конец). (ДНК-полимераза может присоединять новые нуклеотиды только к 3’-концу.)
Две полинуклеотидные цепи соединяются друг с другом слабыми водородными связями между азотистыми основаниями. Соблюдаются 2 правила:
принцип комплементарности: напротив аденина всегда стоит тимин, напротив цитозина – гуанин (они подходят друг другу по форме и числу водородных связей – между А и Г две связи, между Ц и Г – 3).
принцип антипараллельности: там, где у одной полинуклеотидной цепи 5’-конец, у другой – 3’-конец, и наоборот.
Получается двойная цепь ДНК.
Она скручивается в двойную спираль , один виток спирали имеет длину 3,4 нм, содержит 10 пар нуклеотидов. Азотистые основания (хранители генетической информации) находятся внутри спирали, защищенные.
Структурная организация молекулы ДНК. Модель Дж.Уотсона и Ф.Крика
В 1950 г. английский физик М.Уилкинс получил рентгенограмму кристаллических волокон ДНК. Она показала, что молекула ДНК имеет определенную структуру, расшифровка которой помогла бы понять механизм функционирования ДНК. Рентгенограммы, полученные не на кристаллических волокнах ДНК, а на менее упорядоченных агрегатах, которые образуются при более высокой влажности, позволили Розалинд Франклин, коллеге М. Уилкинса, увидеть четкий крестообразный рисунок - опознавательный знак двойной спирали. Стало известно также, что нуклеотиды расположены друг от друга на расстоянии 0,34 нм, а на один виток спирали их приходится 10. Диаметр молекулы ДНК составляет около 2 нм. Из рентгеноструктурных данных, однако, было не ясно, каким образом цепи удерживаются вместе в молекулах ДНК.
Картина полностью прояснилась в 1953 г., когда американский биохимик Дж. Уотсон и английский физик Ф. Крик, исследуя структуру молекулы ДНК, пришли к выводу, что сахарофосфатный остов находится на периферии молекулы ДНК, а пуриновые и пиримидиновые основания - в середине. Причем последние ориентированы таким образом, что между основаниями из противоположных Цепей могут образоваться водородные связи. Из построенной ими модели выявилось, что какой-либо пурин в одной цепи всегда связан водородными связями с одним из пиримидинов в другой цепи. Такие пары имеют одинаковый размер по всей длине молекулы. Не менее важно то, что аденин может спариваться лишь с тимином, а гуанин только с с цитозином. При этом между аденином и тимином образуются две водородные связи, а между гуанином и цитозином – три.
Свойства и функции ДНК.
Хранение наследственной информации (генетический код – способ записи ген.информации о последовательности аминокислот в белке с помощью нуклеотидов (Гамов)
Передача (репликация/удвоение)
Реализация (транскрипция)
Ауторепродукция ДНК. Репликон и его функционирование.
Процесс самовоспроизведения молекул нуклеиновых кислот, сопровождающийся передачей по наследству (от клетки к клетке) точных копий генетической информации; осуществляется с участием набора специфических ферментов (геликаза, контролирующая расплетание молекулы ДНК, ДНК-полимеразы, ДНК-лигаза), проходит по полуконсервативному типу с образованием репликативной вилки; на одной из цепей синтез комплементарной цепи непрерывен, а на другой происходит за счет образования фрагментов Дказаки. Высокоточный процесс, частота ошибок при котором не превышает 10 -9 ; у эукариот может происходить сразу в нескольких точках одной молекулы ДНК; скорость у эукариот около 100, а у бактерий - около 1000 нуклеотидов в сек.
Репликон - единица процесса репликации участка генома, который находится под контролем одной точки инициации (начала) репликации. Термин предложен Ф. Жакобом и С. Бреннером в 1963 году. Геном прокариот представляет собой, как правило, один репликон. От точки инициации репликация идёт в обе стороны, в некоторых случаях с неравной скоростью. У эукариот геном состоит из многих (часто до неск. десятков тысяч) репликонов.
Генетический код, его свойства.
Генетический код – способ записи генетической инофрмации о последовательности аминокислот в белке с помощью нуклеотидов. Открытие ген. Кода принадлежит Георгию Гамову. 1954год.
Триплетность - значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон).
Непрерывность - между триплетами нет знаков препинания, то есть информация считывается непрерывно.
Неперекрываемость - один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов (не соблюдается для некоторых перекрывающихся генов вирусов, митохондрий и бактерий, которые кодируют несколько белков, считывающихся со сдвигом рамки).
Однозначность (специфичность) - определённый кодон соответствует только одной аминокислоте (однако, кодон UGA у Euplotes crassus кодирует две аминокислоты - цистеин и селеноцистеин)
Вырожденность (избыточность) - одной и той же аминокислоте может соответствовать несколько кодонов.
Универсальность - генетический код работает одинаково в организмах разного уровня сложности - от вирусов до человека (на этом основаны методы генной инженерии; есть ряд исключений, показанный в таблице раздела «Вариации стандартного генетического кода» ниже).
Помехоустойчивость - мутации замен нуклеотидов, не приводящие к смене класса кодируемой аминокислоты, называют консервативными ; мутации замен нуклеотидов, приводящие к смене класса кодируемой аминокислоты, называют радикальными .
Понятие о гене. Свойства Гена.
Ген - структурная и функциональная единица наследственности живых организмов. Ген представляет собой последовательность ДНК, задающую последовательность определённого полипептида либо функциональной РНК. Гены определяют наследственные признаки организмов, передающиеся от родителейпотомству при размножении. При этом некоторые органеллы (митохондрии, пластиды) имеют собственную ДНК, не входящую в геном организма, которая определяет их признаки.
(Термин введен в 1909 году датским ботаником Вильгельмом Йогансеном)
дискретность - несмешиваемость генов;
стабильность - способность сохранять структуру;
лабильность - способность многократно мутировать;
множественный аллелизм - многие гены существуют в популяции во множестве молекулярных форм;
аллельность - в генотипе диплоидных организмов только две формы гена;
специфичность - каждый ген кодирует свой признак;
плейотропия - множественный эффект гена;
экспрессивность - степень выраженности гена в признаке;
пенетрантность - частота проявления гена в фенотипе;
амплификация - увеличение количества копий гена.
Особенности организации генома эукариот.
Геном эукариот:
большое число генов,
большее количество ДНК,
в хромосомах имеется очень сложная система контроля активности генов во времени и пространстве, связанная с дифференциацией клеток и тканей в онтогенезе организма.
Количество ДНК в хромосомах велико и возрастает по мере усложнения организмов. Для эукариот также характернаизбыточность генов. Так, у человека геном содержит число нуклеотидных пар, достаточное для образования более 2 млн. структурных генов, в то время как у человека имеется по данным 2000 года 31 тыс. всех генов.
Больше половины гаплоидного набора генома эукариотов составляют уникальные гены, представленные лишь по одному разу. У человека таких уникальных генов - 64%, у теленка - 55%, у дрозофилы - 70%.
Классы нуклеотидных последовательностей в ДНК эукариот, их характеристика, свойства и биологичесок значение.
Нуклеотидные последовательности в геноме эукариот
В конце 60-х годов работами американских ученых Р. Бриттена, Э. Дэвидсона и других была открыта фундаментальная особенность молекулярной структуры генома эукариот – нуклеотидные последовательности разной степени повторяемости. Это открытие было сделано с помощью молекулярно-биологического метода изучения кинетики ренатурации денатурированной ДНК. Различают следующие фракции в геноме эукариот.
1. Уникальные, т.е. последовательности, представленные в одном экземпляре или немногими копиями. Как правило, это цистроны – структурные гены, кодирующие белки.
2. Низкочастотные повторы – последовательности, повторяющиеся десятки раз.
3. Промежуточные, или среднечастотные, повторы – последовательности, повторяющиеся сотни и тысячи раз. К ним относятся гены рРНК (у человека 200 на гаплоидный набор, у мыши – 100, у кошки – 1000, у рыб и цветковых растений – тысячи), тРНК, гены рибосомных белков и белков-гистонов.
4. Высокочастотные повторы, число которых достигает 10 миллионов (на геном). Это короткие (~ 10 пн) некодирующие последовательности, которые входят в состав прицентромерного гетерохроматина.
Уровни организации генома эукариот.
Химический и структурный состав хромосом.
Молекулярно-биологические исследования позволили получить представление не только о химической структуре хромосом, но также и об их надмолекулярной организации и особенностях функционирования. В настоящее время известно, что хромосомы представляют собой нуклеопротеидные образования, состоящие из ДНК и белка. Кроме того, в хромосомах присутствует некоторое количество РНК, образующейся при транскрипции, и ионы Са+ и Mg+. Каждая хроматида, а в промежутке времени анафаза- S -период интерфазы и хромосома, содержит одну молекулу ДНК, которая определяет все функции хромосомы, связанные с хранением наследственной информации, её передачей и реализацией. Молекула ДНК в хромосомах тесно связана с двумя классами белков- гистонами (основные белки) и негистонами (кислые белки). Гистоны - это небольшие по величине белки с высоким содержанием заряженных аминокислот (лизина и аргинина). Суммарный положительный заряд позволяет гистонам связываться с ДНК независимо от нуклеотидного состава. Им принадлежит в основном структурная функция. Это очень стабильные белки, молекулы которых могут сохраняться в течение всей жизни клетки. В эукариотической клетке присутствуют 5 типов гистонов, которые распределяются на две основные группы: первая группа (их обозначают как Н2А, Н2В, НЗ, Н4), отвечает за формирование специфических дезоксирибонуклеопротеидных комплексов - нуклеосом. Вторая группа гистонов (HI) располагается между нуклеосомами и фиксирует укладку нуклеосомной цепи в более высокий уровень структурной организации (супернуклеосомную нить). Среди гистоновых белков, кроме структурных, встречаются такие, которые способны ограничивать доступность ДНК для ДНК - связывающих регуляторных белков и тем самым участвовать в регуляции активности генов. Негистоновые белки весьма разнообразны. Число их фракций превышает 100. Они присутствуют в меньших количествах в хромосомах в сравнении с гистонами и выполняют в основном регуляторную функцию. Участвуют в регуляции транскрипционной активности генов, в обеспечении редупликации и репарации ДНК. Большинство негистоновых белков хроматина присутствуют в клетках в небольшом количестве (минорные) - это регуляторные белки, узнающие специфические последовательности ДНК и связывающиеся с ними. Они вовлечены во многие генетические процессы, но известно о них пока что немного. Количественно преобладают негистоновые белки (мажорные), высокоподвижные, относительно малого размера, с большим электрическим зарядом - они всегда соединяются с нуклеосомами, содержащими активные гены. Кроме того, в группу негистоновых белков входит много ферментов.
Уровни упаковки наследственного материала у эукариот.
Таким образом, уровни упаковки ДНК следующие:
1) Нуклеосомный (2,5 оборота двуспиральной ДНК вокруг восьми молекул гистоновых белков).
2) Супернуклеосомный - хроматиновая спираль (хромонема).
3) Хроматидный - спирализованная хромонема.
4) Хромосома - четвертая степень сперализации ДНК.
В интерфазном ядре хромосомы деконденсированы и представлены хроматином. Деспирализованный участок, содержащий гены, называется эухроматин (разрыхленный, волокнистый хроматин). Это необходимое условие для транскрипции. Во время покоя между делениями определенные участки хромосом и целые хромосомы остаются компактными.
Эти спирализованные, сильно окрашивающиеся участки, называются гетерохроматином. Они неактивны в отношении транскрипции. Различают факультативный и конститутивный гетерохроматин.
Факультативный гетерохроматин информативен, т.к. содержит гены и может переходить в эухроматин. Из двух гомологичных хромосом одна может гетерохроматической. Конститутивный гетерохроматин всегда гетерохроматичен, неиформативен (не содержит генов) и поэтому всегда неактивен в отношении транскрипции.
Хромосомная ДНК состоит из более 10 8 пар оснований, из которых образуется информативные блоки - гены, расположенные линейно. На их долю приходится до 25% ДНК. Ген - функциональная единица ДНК, содержащая информацию для синтеза полипептидов, или всех РНК. Между генами находятся спейсеры - неинформативные отрезки ДНК разной длины. Избыточные гены представлены большим числом - 10 4 идентичных копий. Примером являются гены для т-РНК, р-РНК, гистонов. В ДНК встречаются последовательности одних и тех же нуклеотидов. Они могут быть умеренно повторяющимися и высоко повторяющимися последовательностями. Умеренно повторяющиеся последовательности достигают 300 пар нуклеотидов с повторениями 10 2 - 10 4 и представляют чаще всего спейсеры, избыточные гены.
Высокоповторяющиеся последовательности (10 5 - 10 6) образуют конститутивный гетерохроматин. Около 75% всего хроматина не участвует в транскрипции, он приходится на высокоповторяющиеся последовательности и нетранскрибируемые спейсеры.
Морфологические особенности метафазной хромосомы.
Митотическая суперкомпактизация хроматина делает возможным изучение внешнего вида хромосом с помощью световой микроскопии. В первой половине митоза они состоят из двух хроматид, соединенных между собой в области первичной перетяжки (центромеры или кинетохора ) особым образом организованного участка хромосомы, общего для обеих сестринских хроматид. Во второй половине митоза происходит отделение хроматид друг от друга. Из них образуются однонитчатые дочерние хромосомы, распределяющиеся между дочерними клетками.
В зависимости от места положения центромеры и длины плеч, расположенных по обе стороны от нее, различают несколько форм хромосом: равноплечие, или метацентрические (с центромерой посередине), неравноплечие, или субметацентрические (с центромерой, сдвинутой к одному из концов), палочковидные, или акроцентрические (с центромерой, расположенной практически на конце хромосомы), и точковые -очень небольшие, форму которых трудно определить (рис. 3.52). При рутинных методах окраски хромосом они различаются по форме и соотносительным размерам. При использовании методик дифференциальной окраски выявляется неодинаковая флуоресценция или распределение красителя по длине хромосомы, строго специфические для каждой отдельной хромосомы и ее гомолога (рис. 3.53).
Таким образом, каждая хромосома индивидуальна не только по заключенному в ней набору генов, но и по морфологии и характеру дифференциального окрашивания.
Эу- и гетерохроматин, их биологическое значение.
Некоторые хромосомы во время клеточного деления выглядят конденсированными и интенсивно окрашенными. Такие различия были названы гетеропикнозом. Для обозначения районов хромосом, демонстрирующих положительный гетеропикноз на всех стадиях митотического цикла был предложен термин «гетерохроматин». Различают эухроматин - основную часть митотических хромосом, которая претерпевает обычный цикл компактизации декомпактизации во время митоза, и гетерохроматин - участки хромосом, постоянно находящиеся в компактном состоянии.
У большинства видов эукариот хромосомы содержат как эу-, так и гетерохроматиновые участки, причем последние составляют значительную часть генома. Гетерохроматин располагается в прицентромерных, иногда в прителомерных областях. Обнаружены гетерохроматиновые участки в эухроматиновых плечах хромосом. Они выглядят как вкрапления (интеркаляции) гетерохроматина в эухроматин. Такой гетерохроматин называют интеркалярным. Компактизация хроматина. Эухроматин и гетерохроматин различаются по циклам компактизации. Эухр. проходит полный цикл компактизации-декомпактизации от интерфазы до интерфазы, гетеро. сохраняет состояние относительной компактности. Дифференциальная окрашиваемость. Разные участки гетерохроматина окрашиваются разными красителями, некоторые районы - каким-то одним, другие - несколькими. Применяя различные красители и используя хромосомные перестройки, разрывающие гетерохроматиновые районы, у дрозофилы удалось охарактеризовать много небольших районов, где сродство к окраскам отлично от соседних участках.
Понятие о кариотипе (определение).Обща характеристика кариотипа человека.
Кариотип - диплоидный набор хромосом, свойственный соматическим клеткам организмов данного вида, являющийся видоспецифическим признаком и характеризующийся определенным числом, строением и генетическим составом хромосом.
Если число хромосом в гаплоидном наборе половых клеток обозначить п, то общая формула кариотипа будет выглядеть как 2п, где значение п различно у разных видов. Являясь видовой характеристикой организмов, кариотип может отличаться у отдельных особей некоторыми частными особенностями. Например, у представителей разного пола, имеются в основном одинаковые пары хромосом (аутосомы ), но их кариотипы отличаются по одной паре хромосом (гетерохромосомы, или половые хромосомы ). Иногда эти различия состоят в разном количестве гетерохромосом у самок и самцов (XX или ХО). Чаще различия касаются строения половых хромосом, обозначаемых разными буквами -X и Y (XX или XY).
Каждый вид хромосом в кариотипе, содержащий определенный комплекс генов, представлен двумя гомологами, унаследованными от родителей с их половыми клетками. Двойной набор генов, заключенный в кариотипе,- генотип - это уникальное сочетание парных аллелей генома. В генотипе содержится программа развития конкретной особи.
Денверская (1960) и Парижская (1971) классификация хромосом человека: основные принципы и сущность.
Денверская и Парижская классификация хромосом Хромосомы подразделяются на аутосомы (соматических клеток) и гетерохромосомы (половых клеток). По предложению Левитского (1924) диплоидный набор соматических хромосом клетки был назван кариотипом. Он характеризуется числом, формой, размерами хромосом. Для описания хромосом кариотипа по предложению С.Г. Навашина их располагают в виде идиограммы - систематизированного кариотипа. В 1960 году была предложена Денверская международная классификация хромосом, где хромосомы классифицированы по величине и расположению центромеры. В кариотипе соматической клетки человека различают 22 пары аутосом и пару половых хромосом. Набор хромосом в соматических клетках называют диплоидным , а в половых клетках - гаплоидным (он равен половине набора аутосом). В идиограмме кариотипа человека хромосомы делят на 7 групп, в зависимости от их размеров и формы. 1 - 1-3 крупные метацентрические. 2 - 4-5 крупные субметацентрические. 3 - 6-12 и Х-хромосома средние метацентрические. 4 - 13-15 средние акроцентрические. 5 - 16-18 относительно малые мета-субметацентрические. 6 - 19-20 малые метацентрические. 7 - 21-22 и Y-хромосома наиболее малые акроцентрические. Согласно Парижской классификации хромосомы разделены на группы по их размерам и форме, а также линейной дифференцировке.